|
Добавлено 26-06-2010
|
|
Эволюция кариотипов муравьёв с обзором числа хромосом
|
|
Pedro LORITE & Teresa PALOMEQUE
E-mail:
plorite@ujaen.es
Dr. Pedro Lorite (contact author) & Dra. Teresa Palomeque, Departamento de Biologia Experimental, Area de Genetica, Universidad de Jaen, 23071 Jain, Spain. plorite@ujaen.es
Karyotype evolution in ants (Hymenoptera: Formicidae) with a review of the known ant chromosome numbers
|
|

"Myrmecologische Nachrichten"
("Myrmecological News")
2010, Volume 13,
Pages 89-102
(Wien, апрель 2010)
|
|
|
Исследована эволюция кариотипов муравьёв с полным обзором всех известных хромосомных наборов семейства Formicidae. В приложении к статье приводится подробный список 750 хромосомных наборов.
Муравьи (Ants; Hymenoptera: Formicidae) представляют собой весьма разнообразную группу насекомых с боле чем 12 000 описанными видами. Подобно другим Перепончатокрылым насекомым (осам, пчёлам, наездникам, пилильщикам), они демонстрируют систему гаплодиплоидии, когда из оплодотворённых яиц развиваются диплоидные самки (матки и рабочие особи), а из неоплодотворённых яиц появляются только гаплоидные самцы. Большое количество видов цитологически исследованы. Хромосомные наборы в настоящее время известны у более чем 750 видов муравьёв. Все эти данные суммированы в этой статье. Formicidae это одна из наиболее изменчивых в отношении хромосомного набора групп организмов. Гаплоидный набор у муравьёв варьирует в широких пределах у разных видов от n = 1 до n = 60. This chromosome diversity suggests that karyotype modifications have accompanied ant diversification. Karyotype evolution has followed chromosome-mutation processes able to change not only chromosome number but also chromosome morphology. We review the different chromosome mutations observed in ants and die possible role of such mutations in karyotype evolution in these insects, and we examine die hypotheses proposed to explain how this karyotype evolution may have occurred. Among chromosome rearrangements. Robertsonian centric fusions and fissions, besides inversions and translocations, seem to be the mam processes mat generate changes in ant karyotypes. Other processes altering chromosome numbers, such as polyploidy or aneuploidy. do not appear to be important in ant evolution Ant subfamilies present different levels of variation in relation to chromosome number.
Наибольшая вариация хромосомного набора обнаружена в примитивных подсемействах, таких как Ponerinae (n = 3 - 60) и Myrmeciinae (n = 1 - 47), в то время как у менее примитивных подсемействах хромосомные наборы менее вариабельны, как например, у Dolichoderinae (n = 5 -16), Formicinae (n = 8 -28) и Myrmicinae (n = 4 - 35). По другим подсемействам данные отрывочны но имеются для общего сравнения. Primitive ants present not only the highest range of variation in chromosome number but also the most complex chromosome polymorphisms. In contrast, less primitive genera show lower variation in chromosome number, and generally only simple polymorphisms have been detected. We conclude with an outlook on future research avenues.
Keywords: Formicidae. cytogenetics, chromosome number, karyotype, evolution, genetics, haplo-diploid. review.
См. ниже частичный перевод статьи, ЧИСЛО ХРОМОСОМ и список литературы.
|
Formicidae
Хромосомные гаплоидные наборы муравьёв Formicidae
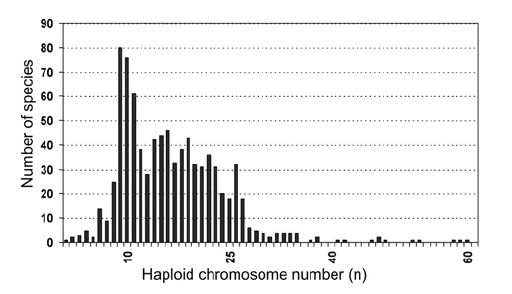
Fig.1. Хромосомные гаплоидные наборы муравьёв Formicidae
(по: ©Lorite & Palomeque, 2010)
|
См. ниже частичный перевод статьи, число хромосом и список литературы.
|
| |
|
|
|
| |
ХРОМОСОМЫ МУРАВЬЁВ: ЭВОЛЮЦИЯ КАРИОТИПОВ
ВВЕДЕНИЕ
Муравьи (Hymenoptera: Formicidae) это одна из наиболее отличительных и хорошо исследованных групп насекомых, которая представляется хорошо обособленным естественным семейством. Being social msects. ants live in colonies, with two female castes (workers and queens) and males which present morphological and genetic differences. The main genetic differences are that males are haploid while queens and workers are diploid. Haplo-diploidy or male haploidy is the mam genetic characteristic of the order Hymenoptera.
Из более чем 12,000 описанных видов муравьёв (Bolton & al. 2007), многие были цитогенетически исследованы, хотя большинство из таких работ касались только числа хромосом и кариотипа. Эти исследования показали, что муравьи принадлежат к эукариотам с самым варьирующим чилом хромосом у разных видов, в пределах от n (гаплоидное число хромосом) = 1 до n = 60. This wide variation could be due to the evolution of this old family, which appears to have diversified more than 100 million years ago (Brady & al. 2006, Moreau & al. 2006, MOREAU 2009).
В муравьиной цитогенетике основополагающими работами были труды двух учёных, это Hirotami T. Imai и Rossiter H. Crozier, которые, работая совместно, проанализировали большинство кариотипов муравьёв. Crozier издал монографию в 1975, где сделал обзор цитогенетики всех Hymenoptera (Crozier 1975). В это работе собраны все данные по числу хромосом на 1975 год. В 1977 году, Imai & al. опубликовали статью о муравьях Австралии, где суммировали данные о кариотипах 150 видов. Later, another key paper was published on Indian ants with the data of 94 species (Imai & al. 1984). These works are important not only for the large number of species analysed but also because chromosome polymorphism m ants is analysed and a hypothesis for chromosome evolution in tins group, the "minmium-interaction theory", is given for the first tune. Tins theory seeks to establish the possible mechanisms governing ant karyotype evolution (Imai & al. 1986, 1988a).
Karyological analysis lias proven useful to determine the karyotypic relationships between related species and then evolution (Imai 1971, Palomeque & al. 1998, Loiselle & al. 1990, Palomeque & al. 1993b). Kaiyological analysis has also proven useful to establish and characterise new species (Imai & al. 1994). since speciation can occur with a modification in chromosome number or with chances in chromosome morphology.
Since Crozier's work, several reviews on chromosome numbers have been made. These studies, however, did not include a large number of species and are limited to countries, regions, subfamilies, tribes or genera (Hauschteck-Jungen & JUNGEN 1976, ROSENGREN & al. 1980, GONI & al. 1983, Hauschteck-Jungen & Jungen 1983, Fischer 1987, Loiselle & al. 1990, Lorjte & al. 1998a, b, 2000, Mariano & al. 2003, 2006, 2007). Таким образом, если в 1975 году были цитогенетически исследованы хромосомы примерно 200 видов муравьёв (Crozier 1975), то на сегодня хромосомные наборы известны более чем у 750 видов муравьёв.
|
Formicidae по подсемействам
Хромосомные гаплоидные наборы муравьёв разных подсемейств
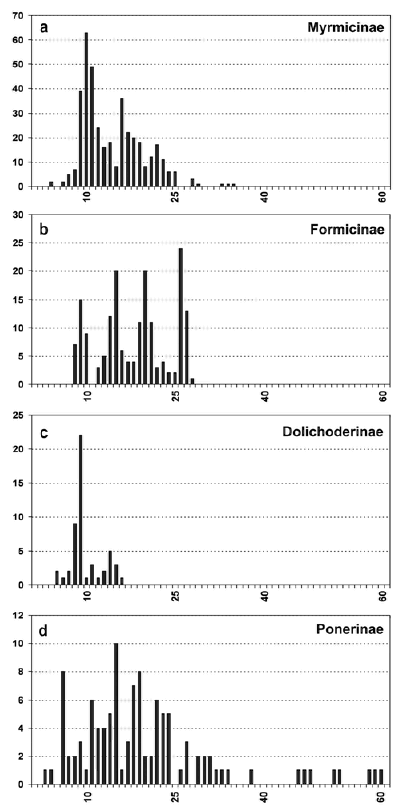
Fig.2. Хромосомные гаплоидные наборы муравьёв разных подсемейств
(по: ©Lorite & Palomeque, 2010)
|
Число хромосом у Formicidae
Большинство данных по числу хромосом, рассматриваемых в данной статье, продятся в приложении к статье (Appendix). Formicidae are highly variable in terms of chromosome number. Самое низкое число хромосом обнаружено Myrmecia crosiandi, n = 1 (CROSLAND & CROZIER 1936, TAYLOR 1991), а самое высокое у Dinoponera lucida, n = 60 (MARIANO & al. 2008). На Figure 1 показано распределение гаплоидных хромосомных наборов среди 750 видов муравьёв. Это распределение в основном напоминает такое же, приведённое ещё ранее IMAI & al. (1988a) на основе 500 видов муравьёв.
Большинство кариотипированных видов принадлежит к подсемействам Myrmicinae, Formicinae, Dolichoderinae и Ponerinae. У Myrmicinae с более чем 400 кариотипированными видами, от n = 4 до n = 35. Гистограмма Myrmicinae сходна с данными по всему семейству муравьёв (Fig. 1, Fig. 2a) как и со старой гисторгаммой, сообщенной IMAI & al. (1988a).
В подсемействе Formicinae, с более чем 400 кариотипированными видами, число хромосом варьирует от n = 8 до n = 28 (Fig. 2b). The histogram differentiates four groups of species with different "modal" values (i.e., groups that differ in the values mat are most frequent). Виды с n = 8 - 10 are представлены главным образом родами Camponotus, Lepisiota, и Plagiolepis. Группа с n = 14 - 15 включает главным образом представителей родов Lasius, Paratrechina и некоторых Camponotus. Группа с n= 19-21 также включает представителей рода Camponotus и большинство видов рода Polyrhachis. Последняя группа с n = 26 - 27, представлена Formica, Polyergus, и Cataglyphis.
Известные на сегодня числа хромосом для подсемейства Dolichoderinae менее вариабельны, чем в других подсемействах, n = 5 - 16 (Fig. 2c). В этом подсемействе около 50 кариотипированных видов. The modal value is n = 9. This modal value is due to the fact that this chromosome number is common in most of Iridomyrmex species, although n = 9 also occurs in other genera.
Около 100 видов подсемейства Ponerinae также были кариотипирован.В этом подсемейства наблюдается очень большая вариация хромомосомных наборов, n = 3 - 60, with a modal value of n = 15 (Fig. 2d). Это подсемейство включает один из наиболе вариабельных по числу хромосом род муравьёв, а именно Pachycondyla, с n = 6 до n = 52 (MARIANO & al. 2007).
В подсемействе Myrmeciinae были проанализированы оба его рода (Myrmecia и Nothomyrmecia). Виды рода Myrmecia обладают высочайшей вариабельностью хромомосоного набора, от n = 1 до n = 42. Вид Nothomyrmecia macrops, единственный реликтовый вид рода Nothomyrmecia, имеет набор n = 47 (IMAI & al. 1990). For the remaining ant subfamilies, a low number of species have been karyotyped and some even completely lack cytogenetic data (see Appendix).
|
Морфология хромосом
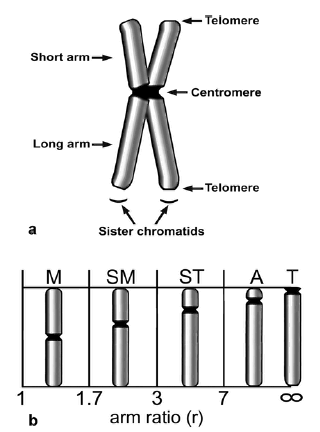
Fig.3. (a) Морфология митотической хромосомы, (b) Классификация хромосом.
Пять различных типов морфологий хромосом разделены по соотношению длин их рук-половинок, то есть длинной и короткой, arm ratio, r. Метацентрические хромосомы (Metacentric) (M) имеют r = 1 to 1.7, субметацентрические (submetacentric) (SM) r = 1.7 - 3, субтелоцентрические (subtelocentric) (ST) r = 3 - 7 и акроцентричекие (acrocentric) (A) r > 7. Телоцентрические хромосомы (Telocentric) (T) имеют ценроимеры в терминальной позиции.
(по: ©Lorite & Palomeque, 2010)
|
Mechanisms of chromosomal changes found in Formicidae
Formicidae
... ... ...
Chromosome rearrangements
Formicidae
... ... ...
Hypotheses of karyotype evolution in Formicidae
Formicidae is one of the insect groups with the greatest variation in chromosome number. It is tentatively assumed that species differentiation has been accompanied by changes in the karyotype.
... ... ...
Karyotype evolution and speciation
Как было сказано выше, chromosome rearrangements could be involved in speciation, since they reduce the fitness of structural hybrids due to the deleterious effect in the meiosis of the heterozygous individuals. Therefore, chromosome mutations could be involved in speciation. as they have the potential to generate postulating isolation mechanisms. However, the relationships between chromosomal changes and speciation are not always clear and often the result of a chromosomal mutation is only chromosome polymorphism (KING 1987). An important aspect in chromosome evolution in ants is their haploid-diploid nature. Because males are haploid. there is no meiosis for gamete production (PALOMEQUE & al. 1990a). Thus, it is possible that Formicidae are more tolerant of chromosome mutations than are other groups of organisms.
Связь между эволюцией кариотипа и процессом видообразования показана в нескольких родах муравьёв. CROZIER (1970b) found that different chromosomal groups are present in Iridomyrmex and that these karyotypic differences were correlated with morphological differences. Twenty-two years later, SHATTUCK (1992a, b) revised the subfamily Dolichoderinae and divided the former Iridomyrmex genus into seven genera: Anonychomyrma, Doleromyrma, Iridomyrmex s.str., Linepithema, Ochetellus, Papyrius, and Philidris. The majority of species of the former Iridomyrmex genus have a chromosome number of n = 9. but species with n = 6 - 8, 11, and 14 have also been detected. CROZIER (1970b) suggested that the species with n = 14 are not closely related to other Iridomyrmex. SHATTUCK (1992a, b) placed these species into the genus Ochetellns, and considered Old World species with n = 9 to belong to the genus Iridomyrmex s.str. Other Old World species have n = 8. Although both karyotypes could easily be related by Robertsonian processes, morphological differences between these groups of species (CROZIER 1968b) have caused them to be assigned to different genera, such as Anonychomyrma or Papyrius (SHATTUCK 1992a, b). There are also morphological differences between species with n = 8 - 9 from the Old World and species with n = 8 - 9 from the New World (CROZIER 1970b). SHATTUCK (1992b) placed the New World species into the genus Linepithema.
Возможная связь между эволюцией кариотипа и процессом видообразования показана для рода Camponotus. MARIANO & al. (2003), reviewing the cytogenetic data for the genus, reported that karyological differentiation has been clearly involved in the diversification of Camponotus, especially by centric fusions-fissions and inversions. This genus has the second largest number of described species and also contains a large number of karyotyped species. Nearly 70 Camponotus species have been studied and the chromosome number is rather variable, rans-ing from n = 9 to n = 26 (see Appendix). The observed karyotypic variation found in this genus could also be a consequence of its polyphyletic origin (BRADY & al. 2006).
Возможно, лучшим примером применения цитогенетических данных для выяснения взаимоотношения видов внутри рода емеется в роде Myrmecia. В комплексе Myrmecia pilosula complex, диплоидное число хромосом варьирует от 2n = 2 до 2n = 32 (IMAI & al. 1977, CROSLAND & CROZIER 1986). IMAI & al. (1994) defined five different species in this species complex according to their karyotypes. A complex polymorphism has been observed in other species of the same genus, such as Myrmecia piliventris with n = 2 - 4, 32 and 2n = 4, 6, 64 (IMAI & al. 1988a), or Myrmecia fulvipes with n = 6, 2n = 12, 48, 50, 60 (IMAI & al. 1977), so that it is possible that there are several sibling species under these taxa.
Однако, этого не наблюдается у других исследованных родов и there are other ant genera in which species differentiation is not explained by important karyotypic variations. Thus, there are well-diversified genera with a high number of species but without visible changes in their karyotypes. Один из этих родов род Formica. В нём исследованы 32 вида и хромосомный набор у них однообразен, n = 26 - 27 (HAUSCHTECK-JUNGEN & JUNGEN 1976, ROSENGREN & al. 1980, LORITE & al. 1998a). Другие примеры родов с весьма постоянным числом хромосом это Lasis и Pogononyrmex (see Appendix). Консервация кариотипа также наблюдается среди близких родов. Напрмер, MOREAU & al. (2006), используя молекулярную технику, показали, что рода Formica, Polyergus и Cataglyphis очень близкородственны. Хотя только 4 вида исследованы в родах Polyergus и Cataglyphis, их числа как и у Formica, n = 26 - 27 (see Appendix). Один из наиболее кариотипически исследованных родов это Pheidole, который всего включает более 1100 видов муравьёв (BOLTON & al. 2007). Молекулярно-филогенетические исследования явно поддерживают монофилию этого рода (MOREAU 2008), в котором около 75 видов кариотипированы, и 65 имеют n = 9 - 10 и все хромосомы метацентрические. Most species with chromosome numbers different from n = 9 - 10 are unidentified species, and are treated as Pheidole spp. in the publications (GON1 & al. 1982, IMAI & al. 1983, 1984).
ВЫВОДЫ
Цитогенетические исследования Formicidae have determined that changes in the karyotype have accompanied genus and species differentiation. Often when several species of one genus have been karyotyped, a variable chromosome number as well as changes in chromosome morphology have been found. The numerical variations have been brought about mainly by Robertsonian changes of centric fusion or fis-sion. Other processes that have altered chromosome numbers, such as polyploidy or aneuploidy. may not play an important role in the evolution of ants. Robertsonian changes besides inversions and translocations seem to be the main processes that generate changes in ant karyotypes. As commented above, there is an asymmetric distribution of chromosome mutations (IMAI & al. 1986). and thus translocations are more frequent in species with low chromosome numbers while Robertsonian changes take place mainly in species of high chromosome numbers.
Согласно теории minimum-interaction theory, число хромосом у муравьёв в общем имеет тенденцию к увеличению. Однако, несмотря на эту возможную тенденцию, увеличение чисел хромосом, кажется, имеет некоторые пределы. Ant karyotype evolution results in a wide diversity of karyotypes, but probably an optimal range in chromosome number exists. Гаплоидное число хромосом муравьёв варьирует от n = 1 до n = 60, но большинство муравьиных хромосомных наборов лежит в пределах между n = 8 и n = 27 (Fig. 1). This idea is not incompatible with the minimum-interaction theory. In fact. this theory proposes mechanisms by which selection could act against low-chromosome-number karyotypes.
Как было сказано выше, ants with low chromosome numbers have larger chromosomes than species with high chromosome numbers (CROZIER 1975, IMAI & al. 1977, 1988a). Consequently, chromosome number and chromosome size are inversely related. Chromosome size is variable but it has been suggested that there are upper and lower tolerance limits, since beyond certain size limits cell-division processes could falter (see SCHUBERT 2007 for a review). In fact, when large numbers of species of a biological group are analysed, the chromosomal-number distribution is similar to a normal symmetric distribution where the majority of the species show intermediate chromosome numbers. This is the pattern of chromosome-number distribution in mammals (IMAI 1986, PARDO-MANUEL DE VlLLENA & Sa-PIENZA 2001), parasitic Hymenoptera (GOKHMAN 2006), and ants (this paper). Thus, selection against extreme chromosome numbers could make the karyotype less variable if the chromosome number remains within the optimal range. Therefore, high or low chromosome numbers and high chromosome-number variability could be basal features.
У муравьёв, экстремальные числа хромосомных наборов характерны для примитивных групп. Молекулярные филогении подтверждают базальную природу понероидной клады, которая включает подсемейства Ponerinae и Amblyoponerinae, среди других (BRADY & al. 2006, MOREAU & al. 2006). Некоторые данные известны для Amblyoponerinae, но Ponerinae включает виды с высочайшими и низшими хромосомными наборами, такими как у Ponera (n = 3 - 6) и Dinoponera (n = 53 - 60). Подсемейство Ponerinae также включает некоторые рода с крупнейшими разбросами в числе хромосом внутри этих родов, например Pachycondyla (n = 6 - 52) и Platythyrea (n = 9 - 47). В формикоидной кладе, в базальном подсемействе Myrmeciinae есть род с одним из высочаших чисел хромосом (Nothomyrmecia, n = 47) и с нижайшим набором (Myrmecia, n = 1 - 42). It would be informative to increase the cytogenetic data in other subfamilies considered basal as a way to confirm whether the trend indeed is a general one in ants, although current data seem to point in this direction (see Appendix),в менее примитивных подсемействах формикоидной клады, таких как Dolichoderinae, Myrmicinae или Formicinae, хромосомные наборы мене вариабельны. Эти подсемейства включают большинство проанализированных видов (около 50 видов из Dolichoderinae, 100 из Formicinae и более чем 400 из Myrmicinae). Глобальная вариация хромосомных наборах в этих подсемействах менее, чем в некоторых родах Ponerinae илил Myrmicinae (n = 5 -16 в Dolichoderinae, n = 8 - 28 в Formicinae, и n = 4 - 35 в Myrmicinae).
Предполагаемая тенденция к оптимизации кариотипа также наблюдается на уровне видов. Как было уже прокомментировано в этом обзоре, различные хромосомные полиморфизмы были найдены у разных видов от всех проанализированных подсемейств. However, in species belonging to less primitive subfamilies, these polymorphisms are generally less complex and they originated from simple changes which usually generate only two karyotypic forms. Единственное исключение среди мирмицин представляет вид Pheidole noda, который имеет 4 полиморфичных кариотипа (n = 17 - 20), due to fusion and fission processes (IMAI & KUBOTA 1975). Среди других Myrmicinae, например вид Leptothorax muscorum обладает вариабельным числом хромосом (n = 16 - 23) (LOISELLE & al. 1990), хотя авторы доказали, что как минимум 4 разных вида принадлежат к этому таксону.
Наоборот, большинство сложных внутривидовых полиморфизмов было обнаружено у видов из более примитивных родов. Вероятно, лучшие примеры таких видов даёт род Myrmecia (Myrmeciinae). У видов этого рода часто встречаетя вариация кариотипа. These variations are due to Robertsonian polymorphisms of centric fusion and fission, although other types of polymorphisms have been detected, such as pericentromeric inversions, translocation, deletions or changes in the amount of heterochromatin (IMAI & al. 1977, 1988a, 1994, IMAI & TAYLOR 1989, MEYNE & al. 1995). Other complex polymorphisms or wide variation in chromosome number have also been detected in Ponera scabra and Dinopononera lucida (Ponerinae) (IMAI & al. 1988a. MARIANO & al. 2008) and in two species from the genus Rhytidoponera (Ectatomminae) (IMAI &al. l977).
Возможное существование оптимального диапазона в числе хромосом не подразумевает, что кариотипы остаются неизменными. Как был прокомментировано, несколько механизмов изменяют число и морфологию хромосом. Так как перестановки хромосом вообще связываются с уменьшением их числа, их фиксация в популяции кажется маловероятной. WHITE (1978) показал, что 4 фактора лидируют в процессе фиксации хромосомных мутаций: genetic drift, meiotic drive, inbreeding, and a selective advantage of the new karyotype. Some of these processes, if not all, depend heavily on population size, or more specifically on the effective population size, which is related to the number of reproductive individuals. The probability of fixation of a new mutation due to the effect of genetic drift, meiotic drive, and inbreeding is increased in small populations. In social insects, the relative effective population size is much lower than in other organisms since only a few individuals can reproduce per colony. The effective population size depends not only on the number of colonies but also the level of polygyny (CHAPMAN & BOURKE 2001). Hence, the appearance of mutations and their fixation in populations could be random processes and could vary among species or between populations of the same species. These stochastic phenomena result in different evolutionary pathways in ant karyotype evolution as compared to other, non-social organisms. Thus, it is possible to find cases in which speciation has occurred without karyotype changes and others in which species of the same genus present very different karyotypes, as has been described in animal groups such as mammals (IMAI 1983, 1986).
Список 750 хромосомных наборов.
|
| |
ЛИТЕРАТУРА
Список по молекулярно-хромосомной теме
-
Biemon. С. 2008: Genome size evolution: within-species variation in genome size. - Heredity 101: 297-298.
-
Bolton В., Alpert. G., Ward P.S. & Naskrecki P. 2007: Bolton's catalogue of ants of the world 1758-2005. - Harvard University Press. Cambridge, MA, CD-ROM.
-
Brady S.G., Schultz T.R., Fisher B.L. & Ward P.S. 2006: Evaluating alternative hypotheses for the early evolution and diversification of ants. - Proceedinss of the National Academy of Sciences of the USA 103:18172-18177.
-
Crosland M.W.J. & CROZIER, R.H. 1986: Myrmecia pilosula, an ant with only one pair of chromosomes. — Science 231: 1278.
-
Crozier R.H. 1968a: Cytotaxonomic studies on some Australian dohchoderme ants (Hymenoptera: Formicidae). — Caryologia 21:241-259.
-
Crozier R.H. 1968b: Interpopulation karyotype differences in Australian Iridomyrmex of the "delectus" group (Hymenoptera: Formicidae: Dohchoderinae). — Journal of the Australian Entomological Society 7: 25-27.
-
Crozier R.H. 1969: Chromosome number polymorphism in an Australian ponerine ant. - Canadian Journal of Genetics and Cytology 11:333-339.
-
Crozier R.H. 1970: Karyotypes of twenty-one ant species (Hymenoptera; Formicidae), with reviews of the known ant karyotypes. - Canadian Journal of Genetics and Cytology 12: 109-128.
-
Crozier R.H. 1975: Hymenoptera. In: JOHN, B. (Ed.): Animal cytogenetics, vol. 3. Insecta 7. - Gebriider Borntraeger. Berlin. Stuttgart 95 pp.
-
Fischer K. 1987: Karyotypuntersuchungen an selbstandigen und sozialparasitischen Ameisen des Tribus Leptothoracini (Hymenoptera. Formicidae) im Hinblick auf ilire Verwandtschafls-beziehimsen. - PliD thesis. Technical University Darmstadt. 219 pp.
-
Glancey B.M., ROMAIN, M.K.S. & CROZIER, R.H. 1976: Chromosome numbers of the red and the black imported fire ants. Solenopsis itrvicta and S. richteri. - Annals of the Entomological Society of America 69: 469-470.
-
Dowton, M., and Austin, A. D. (1994) Molecular phylogeny of the insect order Hymenoptera: Apocritan relationships. Proc. Natl. Acad. Sci. USA 91, 9911-9915.
-
Gimeno C, Belshaw, R., and Quicke D. L. (1997) Phylogenetic relationships of the Alysiinae/Opiinae (Hymenoptera: Brac-onidae) and the utility of cytochrome b, 16S and 28S D2 rRNA. Insect Mol. Biol. 6, 273-284.
-
GONI В., IMAI H.T., KUBOTA M., KONDO M., YONG H.S. & TSO Y.P. 1982: Chromosome observations of tropical ants in Western Malaysia and Singapore. - Annual Report of the National Institute of Genetics (Japan) 32: 71-73.
-
GONI В., ZOLESSI L.C. De & IMAI H.T. 1983: Karyotypes of thirteen ant species from Uruguay (Hymenoptera. Formicidae). -Caiyologia 36: 363-371.
-
GOODISMAN M.A.D., KOVACS J.L. & HUNT B.H. 2008: Functional genetics and genomics in ants (Hymenoptera: Formicidae): the interplay of genes and social life. — Myrmecological News 11: 107-117.
-
HAUSCHTECK E., 1961: Die Chromosomen von fiinf Ameisen-arten. - Revue Suisse de Zoologie 6S: 218-223.
-
HAUSCHTECK-JUNGEN E. & JUNGEN H. 1976: Ant chromosomes. I. The genus Formica. - Insectes Sociaux 23: 513-524.
-
HAUSCHTECK-JUNGEN E. & JUNGEN H. 1983: Ant chromosomes. II. Karyotypes of Western Palearctic species. - Insectes Sociaux 30: 149-164.
-
HlRAI H., YAMAMOTO M.T., OGURA. K., SATTA Y., YAMADA M.. TAYLOR. R.W. & IMAI. H.T. 1994: Multiplication of 28S rDNA and NOR activity in chromosome evolution among ants of the Myrmecia pilosula species complex. - Chromosoma 103: 171-178.
-
HlRAI H., YAMAMOTO M.T., TAYLOR R.W. & IMAI H.T. 1996: Genomic dispersion of 2SS rDNA during karyotypic evolution in the ant genus Myrmecia (Formicidae). - Chromosoma 105: 190-196.
-
Holldobler В., and Wilson E. О. (1990) The Ants. The Belknap Press of Harvard University Press, Cambridge, Mass.
-
HUNG A.C.F., IMAI H.T. & KUBOTA M. 1972: The chromosomes of nine ant species (Hymenoptera: Formicidae) from Taiwan. Republic of China. - Annals of the Entomological Society of America 65: 1023-1024.
-
HUNG A.C.F., VINSON S.B. & SUMMERLIN J.W. 1974: Male sterility in the red imported fire ant. Solenopsis invicta. - Annals of the Entomoloaical Society of America 67: 909-912.
-
Imai H.T. 1966: The chromosome observation techniques of ants and the chromosomes of Formicinae and Myrmicinae. - Acta Hymenopterologica 2: 119-131.
-
Imai H.T. 1986: Modes of species differentiation and karyotype alteration in ants and mammals. In: IWATSUKI K., RAVEN P.H. & BOCK W.J. (Eds.): Modem aspect of species. - University of Tokyo Press. Tokyo, pp. 87-105.
-
Imai. H.T., BARONI URBAN С., KUBOTA M., SHARMA G.P., XARASIMHANNA M.N., DAS B.C., SHARMA A.K., SHARMA A., DEODIKAR G.B., VAIDYA V.G. & RAJASEKARASETTY, M. R. 1984: Karyological survey of Indian ants. - Japanese Journal of Genetics 59: 1-32.
-
Imai H.T., BROWN W.L., KUBOTA M., YONG H.S. & THO Y.P. 1983: Chromosome observations on tropical ants from Western Malaysia. II. - Annual Report of the National Institute of Genetics (Japan) 34: 66-69.
-
Imai H.T., CROZIER R.H. & TAYLOR R.W. 1977: Karyotype evolution in Australian ants. - Chromosome 59: 341-393.
-
Imai H.T., CROZIER. R.H. & TAYLOR. R.W. 1977: Karyotype evolution in Australian ants. - Chromosome 59: 341-393.
-
Imai H.T. & KUBOTA. M. 1972: Karyological studies of Japanese ants (Hymenoptera: Formicidae). Ill Kaiyotypes of nine species in Ponerinae, Fomricinae and Mymiicinae. - Chromosoma 37: 193-200.
-
Imai H.T. & KUBOTA, M. 1975: Chromosome polymorphism in the ant. Pheidole nodus. - Chromosoma 51: 391-399.
-
Imai H.T., MARUYAMA. Т., GOJOBORI. Y.I. & CROZIER. R.H. 1986: Theoretical bases for karyotype evolution. 1. The minimum-interaction hypothesis. - American Naturalist 128: 900-920.
-
Imai H.T.. SATTA, Y. & TAKAHATA. N. 2001: Integrative study on chromosome evolution of mammals, ants and wasps based on the Minimum Interaction Theory. - Journal of Theoretical Biology 210: 475-497.
-
Imai H.T. & TAYLOR. R.W. 1989: Chromosomal polymorphisms involving telomere fusion, centromeric inactivation and centromere shift in the ant Myrmecia (pilosula) n = 1. - Chromosoma 98: 456-460.
-
Imai H.T., Taylor R.W., Crosland M.W.J. & Crozier. R.H. 1988a: Modes of spontaneous chromosomal mutation and karyotype evolution in ants with reference to the minimum interaction hypothesis. - Japanese Journal of Genetics 6$: 159-185.
-
Imai H.T., TAYLOR, R.W. & CROZIER R.H. 1994: Experimental bases for the minimum interaction theory. I. Chromosome evolution in ants of the Myrmecia pilosula species complex (Hymenoptera: Formicidae: Myrmeciinae). - Japanese Journal of Genetics 69: 137-182.
-
Imai H.T., Taylor. R.W., Crozier R.H., Crosland M.W.L. & Browning, G.P. 1988b: Chromosomal polymorphism in the ant Myrmecia (pilosula) n = 1. — Annual Report of the National Institute of Genetics (Japan) 38: 82-84.
-
Imai H.T., Taylor. R.W., Kubota. M., Ogata. K. & Wada M.Y. 1990: Notes of the remarkable karyology of the primitive ant Nothomyrmecia macrops. and of the related genus Myrmecia (Hymenoptera. Formicidae). -Psyche 97: 133-140.
-
LORITE P., CARRILLO J.A., AGUILAR J.A. & PALOMEQUE T. 2004a: Isolation and characterization of two families of satellite DNA with repetitive units of 135 bp and 2.5 Kb in the ant Monomorium subopacum (Hymenoptera, Formicidae). -Cytogenetic and Genome Research 105: 83-92.
-
LORITE P., CARRILLO J.A.. GARCIA. M.F. & PALOMEQUE T. 2000: Chromosome numbers in Spanish Formicidae. III. Subfamily Myrmicinae (Hymenoptera). - Sociobiology 36: 555-570.
-
LORITE P., CARRILLO J.A. & PALOMEQUE T. 2002a: Conservation of (TTAGG)n telomeric sequences among ants (Hymenoptera, Formicidae). - Journal of Heredity 93: 282-285.
-
LORITE P., CARRILLO J.A., TlNAUT A. & PALOMEQUE T. 2002b: Chromosome numbers in Spanish Formicidae. IV. New data of species from the genera Camponotus, Formica, Lasius, Messor, and Monomorium. - Sociobiology 40: 331-341.
-
LORITE P., CARRILLO J.A., TlNAUT A. & PALOMEQUE T. 2004b: Evolutionary dynamics of satellite DNA in species of the genus Formica (Hymenoptera, Formicidae). - Gene 332: 159-168.
-
LORITE P., Сhica Е. & PALOMEQUE. Т. 1996: G-banding and chromosome condensation in the ant Tapinoma nigerrimum. - Chromosome Research 4: 77-79.
-
LORITE P., Сhica E. & PALOMEQUE. T. 1998a: Nunieros cromosomicos en los Formicidos espaiioles. I. Subfamilia Formicinae. - Boletin de la Real Sociedad Espauola de Historia Natural (Seccion Biologia) 94: 23-31.
-
LORITE P., Сhica E. & PALOMEQUE. T. 1998b: Chromosome numbers in Spanish Formicidae. II. Subfamily Dolichoderinae. -Sociobiology 32: 77-89.
-
Lorite P., Renault S., Rouleux-Bonnin F., Bigot S., Periquet G. & PALOMEQUE. T. 2002c: Genomic organization and transcription of satellite DNA in the ant Aphaenogaster subterranea (Hymenoptera. Formicidae). - Genome 45: 609-616.
-
MARIANO C.S.F., DELABIE. J.H.C., CAMPOS L.A.O. & POMPOLO S.G. 2003: Trends in karyotype evolution in the ant genus Camponotus (Hymenoptera: Formicidae). - Sociobiology 42: 831-839.
-
Mariano C.S.F., Delabie J.H.C., Santos J.R.M. & POMPOLO S.G. 2007: Evolucao cariotipica em Pachycondyla spp. (Ponerinae) Neotropicals. — О Biologico 69. Suplemento 2: 409-412.
-
Mariano C.S.F., Pompolo. S.G., Barros L.A.C., Mariano-Neto, E., CAMPIOLO S. & DELABIE J.H.C 2008: A biogeographical study of the threatened ant Dinoponera lucida EMERY (Hymenoptera: Fomiicidae: Ponerinae) using a cytogenetic approach. - bisect Conservation and Diversity 1: 161-168.
-
Mariano C.S.F., Pompolo S.G., Delabie J.H.C. & Campos L.A.O. 2001: Estudos cariotipicos de algumas especies neotropicais de Campoiwtus MAYR (Hymenoptera. Formicidae). -Revista Brasileira de Entomologia 45: 267-274.
-
MEYNE J.. HIRAI. H. & IMAI. H.T. 1995: FISH analysis of the telomere sequences of bulldog ants (Myrmecia: Fonnicidae). -Chromosoma 104: 14-18.
-
MOREAU C.S. 2008: Unraveling the evolutionary history of the hyperdiverse ant genus Phieidole (Hymenoptera: Formicidae). - Molecular Phylogenetics and Evolution 48: 224-239.
-
MOREAU C.S. 2009: Inferring ant evolution in the age of molecular data (Hymenoptera: Formicidae). - Myrmecological News 12:201-210.
-
Moreau C.S., Bell CD., Vila R., Archibald S.B. & Pierce. N.E. 2006: Phylogeny of the ants: Diversification in the age of angiospenns. - Science 312: 101-104.
-
MURAKAMI. Т., FUJIWARA. A. & YOSHIDA. M.C.1998: Cytogenetics of ten ant species of the tribe Attini (Hymenoptera, Formicidae) in Barro Colorado Island, Panama. - Chromosome Science 2: 135-139.
-
Ohnishi, H., and Yamamoto. M-T. The structure of a single unit of ribosomal RNA gene (rDNA) including intergenic subrepeats in the Australian bulldog ant Myrmecia croslandi (Hymenoptera: Formicidae). Zool. Sci. in press.
-
PALOMEQUE Т., CANO. M.A., CHICA E. & DIAZ DE LA GUARDIA R. 1990a: Spermatogenesis in Tapinoma mgerrimum (Hymenoptera, Formicidae). -Cytobios 62: 71-80.
-
PALOMEQUE Т., CHICA E., CANO. M.A. & DIAZ DE LA GUARDIA R. 1988: Kaiyotypes, C-banding. and chromosomal location of active nucleolar organizing regions in Tapinoma (Hymenoptera, Formicidae). - Genome 30: 277-2S0.
-
PALOMEQUE Т., CHICA E. & DIAZ DE LA GUARDIA. R. 1990b: Kaiyotypes. C-banding. chromosomal location of active nucleolar organizing regions, and B-chromosomes in Lasius niger (Hymenoptera, Formicidae). - Genome 33: 267-272.
-
PALOMEQUE Т., CHICA E. & DIAZ DE LA GUARDIA, R. 1993a: Supernumerary chromosome segments in different genera of Formicidae. - Genetica 90: 17-29.
-
PALOMEQUE Т., CHICA E. & DIAZ DE LA GUARDIA. R. 1993b: Karyotype evolution and chromosomal relationships between several species of the genus ApJiaenogasrey (Hymenoptera, Formicidae). - Caryologia 46: 25-40.
-
PALOMEQUE T. & LORITE. P. 2008: Satellite DNA in insects: a review. -Heredity 100: 564-573.
-
ROSENGREN M., ROSENGREN R. & SODERLUND. V. 1980: Chromosome numbers in the genus Formica with special reference to the taxonomical position of Formica uralensis RUZSK. and Formica truncorum FABR. - Hereditas 92: 321-325.
-
Schmitz, J., and Moritz, R. F. A. (1998) Molecular phylogeny of Vespidae (Hymenoptera) and the evolution of sociality in wasps. Mol. Phylogenet Evol. 9, 183-191.
-
Sullender, B.W. (1998) A molecular phylogeny for the Formicidae. http://www. ruf.rice.edu/-bws/molecular_phylogeny.html
-
Taylor. R. W. (1978) Nottiomyrmecia macrops: a living-fossil ant rediscovered. Science 201, 979-985.
-
Ward. P. S., and Brady, S. G. (2003) Phylogeny and biogeography of the ant subfamily Myrmeciinae (Hymenoptera: Formicidae). Invertebrate Systematics 17, 361-386.
-
Wilson, E. 0. (1971) The Insect Societies. Harvard University Press: Cambridge, MA, USA.
|
| |
| |
|
©2010, Vladislav Krasilnikov (translation & supplement)
|
|
Всякое использование без согласования с автором и без активной гиперссылки на наш сайт преследуется в соответствии с Российским законодательством об охране авторских прав.
|
