

Смотрите
|
|
Образ жизни муравьёв (3)
(по Длусскому и др., 1990 с дополнениями)

(Страница обновлена 12.09.2008)
|
[состав семьи][разделение функций][рабовладение][социальный паразитизм][гнездостроение][питание][кормовой участок][движение тандемом][литература]
|
|
Отвечая на многочисленные просьбы, в том числе студентов и школьников, помочь в написании рефератов и докладов о муравьях (то что они мне прислали оказалось излишне упрощенным и с ошибками) и разместить на сайте соответствующую информацию, я пошел им на встречу. Пока мой собственный авторский раздел со страницами об образе жизни муравьёв (их поведении и составе семьи, о разделении функций и социальном паразитизме, гнездостроении и питании и т.д.) и о строении муравьёв ещё не опубликован, то я предлагаю свои черновики с добавлением выдержек из лучших книг о муравьях (которые теперь, к сожалению, малодоступны в библиотеках и магазинах) и с учетом зарубежной научной периодики.
Материал публикуется на нашем сайте только в образовательных целях, и не может быть использован в коммерческих целях. Гиперссылка обязательна. С научно-популярными книгами ознакомьтесь здесь >>>. Муравьиные журналы здесь >>>.
Раздел дополняется и уточняется каждый месяц. Для вопросов >>>LASIUS@narod.ru
|
Питание и кормовой участок
|

Поймали!
(Фото П.Корзуновича)
Питание
Большинство муравьев или хищники (зоофаги), или падальщики (некрофаги) и собирают всякую малоподвижную добычу, как правило насекомых и других беспозвоночных (пауков, многоножек, аннелид). Чаще же - всего понемногу. То есть большинство из них по своей жизненной форме - зоонекрофаги, с уклонами к явным зоофагам или к падальщикам некрозоофагам. Для питания муравьям необходимы два компонента: белковый и углеводный. Белковой пищей служат различные насекомые, а в качестве углеводной пищи муравьи используют падь тлей, щитовок, червецов, листоблошек, цикад и даже гусениц бабочек, и соки деревьев и внецветковый нектар.
Вся пища, приносимая в гнездо, распределяется между всеми членами семьи. Белковая пища потребляется главным образом личинками, и муравьями, которые выкармливают молодых личинок выделениями специальных желез и кормовыми яйцами (кормовые яйца предназначены только для еды и их откладывают самки и молодые рабочие). Углеводную пищу едят имаго - все взрослые муравьи, которые вышли из куколок.
Но экзотики у муравьёв хватает (например, жнецы, листорезы и грибководы) и об этом позже.
Трофаллаксис (=трофоллаксис).
Жидкая пища распространяется среди насекомых путем трофаллаксиса. Муравей набирает падь в зобик, отделенный от желудка клапаном. Благодаря этому пища в зобике не переваривается. Муравей становится в характерную позу, у него изо рта выделяется капелька жидкости. К нему подходит один или несколько муравьев и эту капельку выпивает. В свою очередь они таким же образом кормят других муравьев и так далее по цепочке корм принесенный одним фуражиром становится достоянием всех, или почти всех обитателей гнезда.
Но есть и другое не менее важное значение трофаллаксиса. Кроме распределения пищи трофаллаксис играет важную роль обратной связи. В пищу подмешивается определенное количество выделений желез и феромонов цариц, личинок и рабочих особей. Благодаря этому муравьи узнают количество приплода в семье, наличие или отсутствие самки и т.д.
Жизненные формы (раздел в подготовке).
Доказано, что все виды муравьев, обитающих в одном биогеоценозе, образуют
коадаптивный комплекс (Длуccкий, 1981), члены которого определенным образом
делят между собой пищевые ресурсы биогеоценоза. В тропических странах питание муравьёв разнообразно, вплоть до экзотичного. Например в Южной Америке муравьи-листорезы разводят в своих гнездах грибы-симбионты.
В качестве субстрата используются листья, которые они срезают с деревьев и переносят в муравейники.
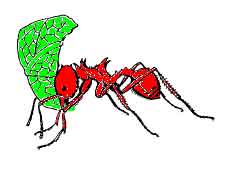
Муравьей-листорез из рода Acromyrmex,
несущий в гнездо часть срезанного листа
|
|
Во влажных местообитаниях, где более или менее выражена ярусность биогеоценозов, муравьи делят среду, главным образом, по вертикали.
Среди жизненных форм муравьёв лесных районов можно выделять следующие группы:
Дендробионты (некоторые Camponotus, Crematogaster
и Leptothorax ) добывают пищу в кронах и на стволах деревьев, хортобионты
- в травостое, герпетобионты ( Tetramorium, Aphaenogaster, Formica и
т.д.) - на поверхности почвы, стратобионты (Myrmica, Ponera, Leptothorax
и др.) в подстилке, геобионты (Lasius flavus) - в почве. Питание этих
муравьев довольно сходно. Все они являются зоонекрофагами, т.е. питаются
малоподвижными насекомыми или их трупами, а также посещают колонии тлей, от
которых они получают падь - экскременты, содержащие большое количество сахара.
Пока мною готовится раздел о жизненных формах у муравьёв ознакомимся с пустынями, о которых хорошо написано у Длусского (1981, 1990). Среди герпетобионтов аридных областей можно выделить 3 главные трофические группы:
1. Карпофаги, или муравьи, питающиеся семенами растений. К этой
группе относятся муравьи-жнецы (Messor) и мелкие карпофаги
(Tetramorium, Pheidole, Monomorium и др.). В пустынях их доля очень велика (до 50-90% биомассы всех муравьёв). Некоторые запасают к концу весны до 1 кг семян, которые они перетирают в муку, смачивают выделениями желез и этой массой питаются.
2. Дневные зоонекрофаги. В эту группу входят представители родов
Cataglyphis, Proformica, Plagiolepis, Formica и др. Источником белка для них являются,
главным образом, трупы насекомых. В наибольшей степени это характерно для рыжих лесных муравьёв и
крупных Cataglyphis. Большинство видов этой группы получает углеводную
пищу, собирая экскременты тлей и других равнокрылых (червецов-кокцид, щитовок; листоблошек-псилид; цикадок), либо сок растений. Многие муравьи не только эксплуатируют, но и охраняют колонии тлей от хищников и паразитов. Все
Formicinae и Dolichoderinae в той или иной степени способны
создавать запасы жидкой пищи, содержащей большое количество сахаров.
3. Ночные хищники. В эту группу входят представители подрода
Tanaemyrmex рода Camponotus. В принципе характер питания
Tanaemyrmex не отличается от такового у других
герпетобионтов-зоонекрофагов, они также собирают трупы насекомых, охотятся на
малоподвижных беспозвоночных на поверхности почвы или собирают падь тлей. Однако
эти насекомые начинают охоту после захода солнца и эаканчивают ее утром.
Большинство зоофагов и зоонекрофагов имеют очень широкий спектр
питания, однако у части видов может наблюдаться определенная специализация. Cerapachys являются специализированными мирмекофагами, т.е. охотятся на других муравьёв.
о видах муравьёв Чувашии
|
|
|
|
Использование кормового участка
Та территория, на которой муравьи из одной семьи собирают пищу носит
название кормового уастка. У наиболее высокоорганизованных муравьев, семьи которых
насчитывают более 100 тыс. особей (Formica s.str., Liometopum и др.),
кормовой участок полностью охраняется от особей своего вида из других семей иди
от других видов со сходной биологией. В этом случае говорят об охраняемой
территория.
Дороги.
У большинства видов муравьев существуют дороги, которые они метят специальными пахучими веществами - феромонами.
Кормовые дороги. Различают дороги кормовые и обменные. Кормовые дороги служат для движения фуражиров и муравьев-охотников, для продвижения к колониям тлей, где муравьи собирают сладкие выделения этих насекомых - падь, служащую основным источником углеводной пищи для муравьев.
Связь муравьев и тлей, называемая трофобиозом, обоюдно выгодна. Это классический пример симбиоза: муравьи получают пищу от тлей и, а тли от них получают защиту от врагов. Кроме того, муравьи переносят тлей на новые побеги и иногда уносят их на зиму в муравейник. Большинство муравьев, идущих от тлей домой имеют светлое раздутое от пади брюшко.
Обменные дороги. Для связи муравейников (семей) служат обменные дороги.
Доказано, что с каждой дорогой связана одна колонна. Колоннами называют определенную группу муравьев, которая живет в одном секторе муравейника и действует на своей ограниченной территории, прилегающем к их дороге. Каждая колонна имеет своих маток и рабочих (строителей, фуражиров, и т.д.).
Неспецифическая активация фуражиров.
У примитивных хищников и некрофагов, например у Роnеrа
coarcata, Cataglyphis emeryi рабочие-фуражиры ведут себя на кормовом участке
совершенно независимо Однако в большинстве случаев у муравьев существует те или
иные способы повышения эффективности использования кормового участка. Простейшая
форма такого взаимодействия - неспецифическая активация фуражиров. Муравей,
обнаруживший пищу (разведчик), возвращается в гнездо и там вызывает возбуждение
других муравьев. Каждый из возбужденных фуражиров направляется туда, где он
обычно добывает корм. В результате, число муравьев на кормовом участке резко
возрастает, что повышает вероятность случайного нахождения того же или другого
источника корма. Такое поведение характерно, например, для фаэтончика
Cataglyphis setipes.
Мобилизация.
Наиболее эффективное средство организации групповой фуражировки -
мобилизация. Под ней понимается комплекс действий разведчика, который приводит к
тому, что другие муравьи (пассивные фуражиры) точно выходят к месту, где
разведчиком был обнаружен корм. Способы мобилизации у разных видов муравьев
могут сильно отличаться. Простейший способ - киноптичеокая мобилизация, был
описан Г. М. Длусским (1981) у пустынного бегунка Cataglyphis foreli при
охоте на термитов. Стоит рабочему этого вида вернуться в гнездо с термитом, как
из него в результате неспецифической активация выходят 10-30 муравьев, которые,
однако, никуда не уходят, а стоят около входа и наблюдают за поведением
разведчика. После второго возвращения разведчика часть из мобилизованных
муравьев уходят в том направлении, откуда тот вернулся. Таким образом,
передается направление к источнику пищи, но не указывается расстояние до нега.
Фактически развитием этого способа является своеобразная
мобилизация пустынных муравьев-жнецов (Захаров, 1972; Длусский,1981). Организацию
групповой фуражировки у них осуществляют активные фуражиры (разведчики),
составляющие всего 3% от их общего числа. Обычно они работает как одиночные
сборщики на своих индивидуальных участках площадью 0,2-0,4 кв.м и все вместе
собирают информацию о ситуации на всем потенциальном кормовом участке. У
муравьев-жнецов рода Messor Г.Длусский выделяет специфический способ мобилизации - дистанционным
наведением. Пассивные фуражиры устремляются вслед за активными в заданном ими направлении, то есть туда, где они недавно нашли много созревших семян. Такой способ мобилизации проявляется в период интенсивной заготовки семян муравьями-жнецами и не дает большой точности, но она и не нужна, т.к. потенциальный источник корма занимает большую площадь. В то же
время он позволяет за короткое время мобилизовать несколько сотен и даже тысяч
фуражиров.
Движение тандемом.
Другие способы мобилизации связаны с точным наведением пассивных
фуражиров на источник пищи простейший из них - движение тандемом. У
Leptothorax разведчик, вернувшись в гнездо, активизирует одного рабочего.
Когда он вновь направляется к корму, мобилизованный муравей идет за ним следом,
постоянно прикасаясь усиками к брюшку и таким образом приходит к корму. Обратно
они возвращаются самостоятельно и приводят еще двух новых муравьев. У
Myrmica, Tetramorium и некоторых Camponotus наблюдается привод
группы. Разведчик, вернувшись в гнездо после обнаружения корма, активизирует
других муравьев при помощи сложного комплекса действий (пробежки, подрагивания
телом, прикосновение усиками и лапками), получившего название осцилляторной
рекрутирующей демонастрации (ОРД), Активизированные рабочие выходят из гнезда и
выстраиваются за разведчиком цепочкой, которую он приводит к корму.
Пахучий след.
Многие муравьи используют для мобилизации пахучий след. Разведчик
от корма к гнезду оставляет пахучий след выделениями специальных желез. У
Formicinae и Ponerinae это ректальные, у Dolichoderinae это
железы Павана, расположенная на нижней стороне брюшка, у Crematogaster -
тарзальные железы и у других Myrmicinae - ядовитая железа (Monomorium,
Tetramorium ) или железа Дюфура (Pheidole, Solenopsis). У Pheidole pallidula
пахучий след сам по себе без дополнительной активизации служит для других
муравьев сигналом к мобилизации. Каждый фуражир, наведший след, оставленный
разведчиком, начинает двигаться по нему в направлении от гнезда и находит корм.
У Lasius, Tapinoma, Crematogaster и многих Monomorium реагируют на
след только фуражиры, предварительно активизированные разведчиком при помощи
ОРД.
Каждый из этих способов мобилизации имеет свои преимущества и
недостатки и зависит от того, какое место в биоценозе занимает вид.
Дистанционное наведение позволяет быстро мобилизовать большое число рабочих, но
при этом точность наведения довольно низка, так что способ хорош для мобилизации
на большие кормовые площади, но непригоден для мобилизации на "точечный"
источник пищи (труп, небольшие группы растений и т.д.). Использование пахучего
следа позволяет на короткое время привести большое число пассивных фуражиров на
"точечный" источник, но этот способ очень инерциален и после исчезновения пищи
муравьи еще долго продолжают ходить по пахучей тропе. Таким образом, он лучше
всего обеспечивает использование "точечных", но довольно крупных источников пищи
(например, трупы крупных насекомых или мелких позвоночных животных). Мобилизация
путем привода группы менее эффективна, но зато группы муравьев могут быстро
переключаться с одного источника на другой, а семья в целом использовать
одновременно несколько источников пищи.
Другой путь повышения эффективности использования кормового
участка связан с совершенствованием структурной организации кормового участка и
основан на вторичном делении его. В простейшем случае, например, у
Cataglyphis, каждый фуражир имеет индивидуальный поисковый участок, на
котором ведет поиск и сбор пищи. Каждый индивидуальный участок при этом
представляет собой сектор в пределах кормового участка семьи.
Сложно устроена территория у некоторых видов Formica, в
частности у рыжих лесных муравьев (группа Formica rufa). У них имеются
колонии - система связанных между собой гнезд, в каждом из которых обитает
отдельная семья. На общей территории колонии имеется сложная система постоянных
дорог, о чем хорошо написано в книгах А.А.Захарова (1972, 1978).
|
|
|
|
ЛИТЕРАТУРА
|
|
Длусский Г. М. "Муравьи рода Формика" - М., 1967. 236 с.
Длусский Г. М., Союнов О. С., Забелин С. И. "Муравьи Туркменистана" - Ашхабад:Ылым, 1990. 275 с.
Длусский Г. М., Букин А. П. "Знакомьтесь, муравьи" - М., 1986. 223 с.
Захаров А. А. "Внутривидовые отношения у муравьев" - М., 1972. 216 с.
Захаров А. А. "Муравей, семья, колония" - М.: Наука. 1978. 144 с.
Кипятков В. Е. "Происхождение общественных насекомых" // "Знание. Сер. Биология" - М., 1985. 64 с.
Кипятков В. Е. "Мир общественных насекомых" - Л., 1991. 396 с.
Халифман И. А. "Муравьи" - М., 1963. 303 с.
Халифман И. А. "Операция "лесные муравьи" - М., 1974.
|
|
|
|

