|
A lattice simulation model to investigate an interaction of herb plant and ant populations
has been developed. The possibility of the ant hill occupying for two species (Lasius
niger and Formica rufa) by puskies (Stellaria holostea) has been analyzed depending on
ant transforming activities. There have been compared the occupying traits of ant-hills and
hill adjacent territories for the investigated species.
***
|
Мирмекологи России >>>
Библиография-РУ >>>
|
|
"ANTS AND FOREST PROTECTION.
Materials of the XIII All-Russian Myrmecological Symposium", Nizhni Novgorod, August 2009
|
|
Введение.
Сообщества муравьев играют полифункциональную роль в есте-
ственных биоценозах. Их вклад в изменение фитоценотической среды
определяется влиянием на физико-химические свойства почвы в процес-
се гнездостроения, нарушении напочвенного покрова, осуществлением
различных этапов семенной инвазии биотопа, участием в динамических
процессах растительности в пределах своих кормовых участков. Суще-
ствует ряд работ (Малоземова, 1970; Бугрова, Пшеницына, 2003; Зрянин
и др., 2004), посвященных выявлению особенностей территориального
размещения трав вблизи муравейников. Однако взаимодействие мура-
вьев и растений по-прежнему остается в числе наиболее актуальных на-
учных задач при разработке основ и методической базы мероприятий
по сохранению полезных лесных муравьев как ценного биологического
ресурса, влияющего на продуктивность и биологическую устойчивость
лесов (Захаров, 2005).
Для изучения взаимоотношений муравьев и растений в данной
работе была разработана и исследована решетчатая имитационная
модель. Она основана на представлении о клеточных автоматах в
обобщении А.С. Комарова с соавторами (Комаров, Паленова, 2001;
Комаров, 2003) для вегетативно-подвижных растений. В модели ис-
пользовался авторский полевой материал сборов 1996–2007 гг. и
литературные данные. Моделировался процесс зарастания муравей-
ников двух видов – черного садового муравья (Lasius niger) и ры-
жего лесного муравья (Formica rufa) – звездчаткой ланцетолистной
(Stellaria holostea).
Расчетная площадка представлялась в виде плоской квадратной ре-
шетки, размер ячейки которой принят равным 4 см. Считаем, что звезд-
чатка ланцетолистная осуществляет вегетативный захват территории
только посредством подземных столонов. В природных условиях в веге-
тативном разрастании принимают участие и надземные побеги, т.к. они
полегают, но их роль в захвате территории мала (Михайлова и др., 2006,
2008). Длина столона не более 10 узлов, в каждом из узлов столона име-
ется почка, из которой возможно прорастание нового побега. При появ-
лении препятствия на пути вегетативного побега, он прекращает свой
рост. Вегетативное разрастание возможно в первый год жизни. Вероят-
ность прорастания почки столона в первый год принята 20%, во все по-
следующие – 40%. Длительность жизни одного элемента популяции – 2
года. Исходное положение сплошного массива звездчатки справа на рас-
стоянии 3 м от муравейника.
Самым сложным при создании любой модели является формали-
зация наблюдаемых явлений. В нашем случае возникла необходимость
описания активности муравейника, его силы. Понятие силы муравей-
ника абстрактно, включает в себя не только численность муравьиной
семьи, но и ее состояние. Хотя для простоты понимания вопроса мож-
но ассоциировать ее с численностью муравьиной семьи. В рассматри-
ваемом случае сила муравейника Ci безразмерная величина. За единицу
силы муравейника принята сила одной гипотетической колонны. Сила
муравейника на i-м шаге описывается следующей формулой:
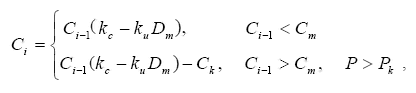
где Ci-1 – сила муравейника на предыдущем шаге, kc = 1,2 и ku – коэффи-
циенты развития и угнетения муравейника соответственно, Dm – доля
купола муравейника, захваченная растениями, Сk = 1 – сила, забираемая
колонной, Рk = 0,5 – вероятность появления колонны, Сm = 2 – сила мура-
вейника, при которой возможно появление колонны.
Купол муравейника располагался в центре расчетной площадки.
Для простоты сравнения диаметр купола принят 1 м для обоих видов.
Способность муравьев освобождать купол муравейника от определен-
ного вида растений характеризуется коэффициентом гибели растений
kdeathhill0 на муравейнике при Ci = 1. При изменении состояния муравей-
ника также меняется коэффициент гибели растений: kdeathhilli = kdeathhill0Ci.
Принято, что угнетающее влияние муравьев на растения одинаково на
всей площади купола. Это влияние за пределами купола у Lasius niger сра-
зу прекращается, а у Formica rufa уменьшается по экспоненте и на границе
выстригаемой территории обнуляется. Ширина кольца выстригаемой
территории 1 м.
Звездчатка ланцетолистная не имеет анатомических приспособле-
ний мирмекохоров, однако, по нашим наблюдениям, муравьи активно
интересуются ее семенами. Отдельные семена звездчатки перемещаются
муравьями F. rufa на расстояние до 7 м, а муравьями Lasius niger до 1,4 м. Счи-
таем, что семена собираются муравьями со всего участка в муравейник.
По пути часть семян теряется, половина принесенных семян выносится и
оставляется рядом с муравейником на расстоянии до 0,5 м. Потери семян
по нашим наблюдениям и литературным данным (Левина, 1957), состав-
ляют 20% от общего количества перенесенных семян. В модели сделано
допущение, что из этих семян всходит на следующий год 3%.
Проведено несколько серий компьютерных экспериментов, в ко-
торых было имитировано развитие растений на куполе муравейников
Formica rufa и Lasius niger и прилегающей к ним территории. Коэффициент смерт-
ности растений на куполе kdeathhill0 изменяли от 0,4 до 1,0. В первом слу-
чае учитывался сбор семян муравьями.
При коэффициенте смертности растений на куполе kdeathhill0 = 0,6 еди-
ничные особи прорастают (рис.). Несмотря на отсутствие выстригаемой
территории у муравейника Lasius niger, характер освоения растениями му-
равейника при тех же значениях kdeathhill0 качественно повторяет картину,
наблюдаемую для Formica rufa. При коэффициенте смертности растений на
куполе kdeathhill0 > 0,6 у растений практически нет шансов появиться на
муравейнике любого из исследуемых видов. Сила муравейника во вре-
мени изменяется «пилообразно» при периодическом отделении отводка
либо бурно растет до появления нового.
При коэффициенте смертности растений на куполе kdeathhill0 < 0,6
муравейник не способен сопротивляться продвижению растений. Рас-
тения, появившиеся на куполе, подавляют развитие семьи. Сила семьи
уменьшается, муравейники Formica rufa и Lasius niger полностью зарастают рас-
тениями и возможно погибают (рис. kdeathhill0 = 0,4).
Серия численных экспериментов без учета семенного размножения
показала, что активный жизнеспособный муравейник Formica rufa недосту-
пен для растений, осваивающих территорию вегетативным способом. В
то же время на муравейник Lasius niger без выстригаемой территории воз-
можно проникновение вегетативно-подвижных растений.
Таким образом, с помощью разработанной модели проведено срав-
нение характера освоения территории муравейников Lasius niger и Formica rufa.
Показано, что возможность захвата территории муравейника растени-
ями зависит от состояния гнезда и биологических особенностей вида
муравьев. Наличие пригнездовой выстригаемой территории вокруг
муравейников F. rufa не является препятствующим фактором для за-
хвата муравейника растениями, чьи семена переносятся муравьями.
Учитывая биологию рассмотренных видов можно сказать, что захват
растениями активного жизнеспособного муравейника видов Lasius niger и
Formica rufa возможен только при коэффициенте смертности растений на ку-
поле kdeathhill0 < 0,6.
|
|
|
|
|
ЛИТЕРАТУРА
-
Бугрова Н.М., Пшеницына Л.Б. Трансформация пространственной структуры тра-
вяного покрова вокруг гнезд Formica aquilonia // УСБ. 2003. Т. 123, № 3. С.
273–277.
-
Захаров А.А. Реальные и мнимые проблемы использования муравьев в биологиче-
ской защите леса // Муравьи и защита леса: Мат. XII Всеросс. мирмекол. симп.
Новосибирск, 2005. C. 224–229.
-
Зрянин В.А., Новоселова Н.А., Петрушова Е.С. Анализ растительных группировок
вокруг муравейников рыжих лесных муравьев (Hymenoptera, Formicidae) //
Поволжский экол. журн. 2004. № 1. С. 48–57.
-
Комаров А.С. Клеточно-автоматные модели сообществ вегетативно-подвижных
растений, учитывающие поливариантность онтогенеза // Математика, ком-
пьютер, образование: Мат. Х Междунар. конф. Ижевск, 2003. Вып. 10, ч. 3. С.
112–124.
-
Комаров А.С., Паленова М.М. Моделирование взаимодействующих популяций
вегетативно-подвижных трав // Бюлл. МОИП. Отд. биол. 2001. Т. 106, № 5. С.
35–41.
-
Малоземова Л.А. О взаимоотношении муравьев с растениями // Экология. 1970.
№ 2. С. 101–103.
-
Михайлова Н.В., Богданова Н.Е, Михайлов А.В. Скорость освоения территории
неморальными видами трав (модельный подход) // Бюлл. МОИП. Отд. биол.
2006. Т. 111, № 1. С. 37–44.
-
Михайлова Н.В., Михайлов A.B., Богданова Н.Е., Комаров А.С., Жукова Л.А. Имита-
ционная модель инвазионной динамики популяций неморальных видов трав
на неоднородной территории // Бюлл. МОИП. Отд. биол. 2008. Т. 113, № 5. С.
68–75.
|
| |
| |
|
©2009, Vladislav Krasilnikov
|
|
Всякое использование без согласования с автором и без активной гиперссылки на наш сайт преследуется в соответствии с Российским законодательством об охране авторских прав.
|
|

