|
Добавлено 12-01-2008
|
|
Филогения, классификация и таксономия муравьев
|
|
Philip S. WARD
E-mail: psward@ucdavis.edu
Department of Entomology, University of California, Davis, CA 95616, USA;
Phylogeny, classification, and species-level taxonomy of ants
(Hymenoptera: Formicidae)*
*In: Zhang, Z.-Q. & Shear, W.A. (Eds). (2007). "Linnaeus Tercentenary: Progress in Invertebrate Taxonomy". Zootaxa,1668, 1-766.
|
|

"Zootaxa", 2007, Vol.1668: pp.549-563.
(December 2007)
|
|
|
Анализ и синтез
АВТОРЕФЕРАТ.
Обзор современного состояния муравьиной систематики. В последние годы построено множество кладограмм основных стволов муравьев с использованием как морфологических, палеонтологических, так и молекулярно-генетических признаков (DNA sequence). Проведен их анализ и синтетический обзор.
[В.К.: замечу здесь, что в последнее время было много сенсационных открытий и выводов по филогении муравьев. Сначала Болтон впервые разделил понероидных муравьев на 6 отдельных подсемейств (B.Bolton, 2003). Это затем было подтверждено и уточнено на молекулярно-генетическом уровне в работах сразу трех независимых коллективов биологов:
американского (Saux et al., 2004),
французского (Astruc et al., 2004) и
японского (Ohnishi et al., 2003).
В в недавней работе (Ouellette et al., 2006) доказано, что и сам понероидный комплекс довольно таки неоднородный и, по-видимому, заслуживает разделения. В структуре всего понероидного комплекса здесь >>>].
Наибольшую сенсацию принесла работа Гарвардского профессора Corrie S. Moreau (отдельное ей спасибо за присланные оттиски её работ), внесшей самые кардинальные за последние полвека изменения в систему подсемейств муравьев. Молекулярные данные с учетом вымерших находок (Moreau, 2006) дают выводы по расхождению двух основных групп муравьев в 140-168 млн.лет (середина юрского периода).
Смотрите ниже частичный перевод этой статьи с большой подробной схемой филогенеза (отдельно 281 Kb) и список литературы.
|
| |
|
|
|
|
взаимоотношения, история и группы подсемейств
подсемейства муравьев (обзор)
систематика муравьев по Б.Болтону >>> (Bolton, 2006): полный список родов, триб и подсемейств
Другие наши обзоры по филогенезу:
муравьи расцвели вместе с цветковыми (Moreau, 2006)
молекулярная филогенетика базального ствола муравьев (rRNA) (Ouellette et al., 2006)
молекулярная филогенетика подсемейств муравьев (rDNA) (Ohnishi, 2003)
подсемейства и ДНК : молекулярная филогенетика
филогенетика Myrmecia (муравьи-бульдоги) и Nothomyrmecia (рибосомальные РНК 18S и 28S)
филогения муравьев-"Дракула" с выводами для всех
Ген abdominal-A и строение стебелька муравьев
Другие наши обзоры по палеонтологической теме:
Вымершие муравьи-бульдоги Myrmeciinae из эоцена Канады >>> (Archibald, 2006)
Новые вымершие, но древнейшие муравьи: Cananeuretus, Myanmyrma, Sphecomyrmodes >>> (Engel, Grimaldi, 2005)
Палеонтологическая история муравьев (Длусский, Расницын)
Древнейшие муравьи из Мела» »-2004: Afropone, Afromyrma, Orapia >>>
Кyromyrma: первая находка в мелу подсемейства Formicinae >>>
Список ~500 видов ископаемых муравьев >>>
Палеонтологическая литература
подробности о систематике муравьев по Длусскому читайте здесь >>>
подробности о подсемействе бульдогов Myrmeciinae читайте здесь >>>
|
Введение
За всю историю было описано около 12 000 видов муравьев, систематика которых была суммаризирована в работах Brown (1955) и B.Bolton (2003). “еперь очевидно, что наибольшее биологическое и таксономическое разнообразие муравьев заключено в внутри Уformicoid cladeФ (схема здесь), которая включает 14 из 20 современных подсемейств и почти 90% видов. являются ли остальные группы (Leptanillinae и понероморфы) анцестральной кладой или градацией (grade) в основании муравьиного ствола остается неясным. Ископаемые находки муравьев из кроны их эволюционного древа уходят назад во времени до 90Ц100 млн.лет. Вымершие виды ствола (sphecomyrmines, armaniines) были также представлены в этот период. Подробнее здесь - Cananeuretus, Myanmyrma, Sphecomyrmodes, (Длусский, Расницын, 2005), Afropone, Afromyrma, Orapia, Kyromyrma.
Молекулярная данные с учетом вымерших находок (Moreau, 2006) дают выводы по расхождению группы (муравьев с учетом других Hymenoptera) показали, что муравьи группы кроны их эволюционного древа возникли ~115-135 млн.лет.назад. Большинство существующих подсемейств муравьев и родов хорошо разграничены морфологически и вероятно монофилетичны, но есть некоторые известные исключения, включая подсемейство Cerapachyinae и несколько больших и двусмысленно разграниченных родов типа Pachycondyla. Несколько триб в больших подсемействах Formicinae и Myrmicinae также представляют искусственный набор. Наконец, в то время как видовая (species level) таксономия некоторых родов муравьев находится в удовлетворительном состоянии, в других родах царит таксономическая анархии, с многочисленными неопределенными разновидностями и многими названиями (именами) сомнительной применимости. Продвижение в этой области систематики муравьев будет требовать объединения индивидуальных усилий, расширение возможностей работы, применение новых технологий, и более глубокого понимания природы разновидностей муравья и различий между ними.
Филогения муравьев
Муравьи это вездесущая и доминантная группа наземных экосистем, играющая ключевую роль в симбиотических отношениях, аэрации почвы и круговороте нутриентов. Они имеют множество ископаемых находок, хотя эволюционная история 12,000 описанных современных видов остается плохо исследованной.
Хотя первые импрессионистские древовидные диаграммы можно найти в ранних работах (Wheeler 1920, Emery 1920, Morley 1938), стартом для дискуссионных современных построений стала классическая муравьиная филогения Brown (1954) опубликованная в журнале Insectes Sociaux.
Браун впервые выделил два ствола-комплекса подсемейств: the Уmyrmecioid
complexФ (Myrmeciinae, Pseudomyrmecinae, Dolichoderinae, Formicinae) и Уponeroid complexФ (Cerapachyinae, Ponerinae, Myrmicinae, Dorylinae, Leptanillinae). Ёта работа (1954), основанная на интуитивной интеграции огромного морфологического материала, имела колоссальное влияние на большинство последующих работ по общей классификации и филогении муравьев.
Новые сенсационные открытие ископаемого мелового муравья Sphecomyrma freyi (Wilson et al.
1967) и переоткрытие живого ископаемого муравья Nothomyrmecia macrops (Taylor 1978), привело к видоизменению схемы Brown (1954).
Почти все выделенные B.Bolton (2003) подсемейства (21) являются монофилетичными, кроме Cerapachyinae. Последние парафилетичны с дориломорфами (Уdoryline sectionФ), от которых более специализированные кочевые муравьи и Leptanilloidinae эволюционировали. Монотипичное подсемейство Apomyrminae ныне размещено внутри Amblyoponinae (Saux et al. 2004). Несмотря на на включение рода Apomyrma в состав Amblyoponinae, возможно это подсемейство парафилетично. Некоторое сходство в поведении и морфологии Leptanillinae и Amblyoponinae (Brown et al. 1971, Bolton 1990b, Masuko 1990, Ward 1994), может говорить о том, что лептаниллины - это сильно модифицированные амблиопонины. молекулярные данные не могут решительно отвергнуть эту гипотезу (Brady et al. 2006).
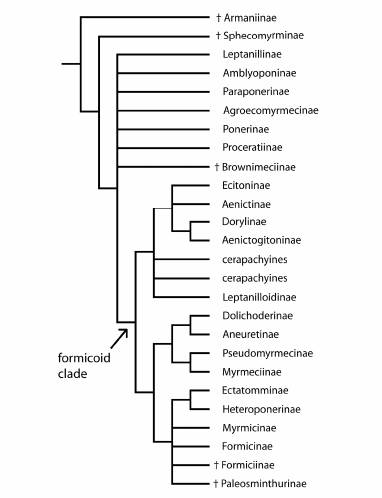
Современные представления филогенеза подсемейств муравьев по молекулярно-генетическим данным (Ward & Brady 2003, Saux et al. 2004, Brady et al. 2006, Moreau et al. 2006, Ouellette et al. 2006). Размещение вымерших таксонов основано по Grimaldi et al. (1997), Dlussky (1999), Ward & Brady (2003) и Bolton (2003).
(©Zootaxa, по: Ward, 2007)
Ископаемые находки и происхождение муравьев
Многочисленные ископаемые находки муравьев и муравьеподобных ос говорят о появлении муравьев примерно в в середине мелового периода (Carpenter 1992, Bolton 2003, Grimaldi & Engel 2005, Perrichot et al. 2007). В мелу находки очень редки (мене 1% от всех насекомых), но позднее (Tertiary) они становятся более массовыми (Grimaldi & Agosti 2000b). Они составляют около 5% о всех насекомых в Балтийском янтаре (эоцен), около 20% насекомых в Florissant shales (ранний олигоцен) и до 36% экземпляров насекомых в доминиканском янтаре (миоцен) (Carpenter 1930, Dlussky & Rasnitsyn 2003).
Дебаты по поводу времени появления первых "настоящих муравьев" (true ant) в ископаемом состоянии (Poinar et al. 1999, 2000, Grimaldi & Agosti 2000a) можно квалифицировать как попытку найти границу между муравьями кроны и ствола муравьиного филогенетического древа (Magallуn 2004). Crown group ants are the clade encompassing the most recent common ancestor of living ants and
their descendants. The stem group is defined more inclusively: all taxa more closely related to ants than to
any other extant organisms. Extinct lineages which lie outside the crown group, but which are considered
more closely related to ants than to any other living aculeate wasps, are members of this more inclusive stem
group. They are here termed stem ants. Thus, phylogenetic analyses place the fossil Sphecomyrma as sister
to the living ants (Grimaldi et al. 1997, Ward & Brady 2003), so it is clearly a stem ant member of the stem
group but not the crown group. Вымершие Armaniinae (иногда в статусе Armaniidae) более отдаленно связаны, но возможно следующие самые близкие известные родственники к существующим муравьям после Sphecomyrminae (Dlussky 1999). Следовательно их можно также рассматривать муравьями стебля филогенетического древа (stem ants). Of course, if the fossil record were sufficiently
detailed we would eventually encounter stem ants with little resemblance to their modern counterparts
because they lack most of the synapomorphies that we associate with ants. This is a dilemma that we do not
currently face, but it has affected arguments about the origin of other fossil-rich groups such as mammals and
flowering plants (de Queiroz & Gauthier 1990, Doyle & Donoghue 1993). Альернативой могло бы стать принятие основанного на апоморфии определения муравьев Formicidae: это все организмы имеющие метаплевральную железу, например. This would place the origin of ants somewhere on the branch below the crown group. Ископаемые сфецомирмины имеют эту железу (Grimaldi et al. 1997), но она отсутствует у Armaniinae, хотя это может зависеть от степени сжатости и сохранности ископаемых останков.
Когда впервые появились ископаемые муравьи из группы кроны эволюционного древа? Ответ на это вопрос зависит от того, сможем ли мы разбросать ископаемые находки по существующим кладам внутри группы кроны (crown group). Большинство третичных муравьев может быть легко помещено в конкретное подсемейство в современные рода (Dlussky & Rasnitsyn 2003). Но то же самое не может быть сказано для меловых остатков муравьев. Ќи один из них, вероятно, не принадлежит к современным родам или трибам, и для большинства из них, даже назначение подсемейства является сомнительным.
Вид Kyromyrma neffi из янтаря Нью-Джерси (Turonian, 90 mya) исключение: он явно имеет ацидопору (Grimaldi & Agosti 2000b), признак характерный только для Formicinae, что делает Kyromyrma более близким родом к этой кладе, чем к какому-либо другому подсемейству. Using relaxed clock divergence dating Brady et al. (2006) obtained an estimated age for crown group
Formicinae of ~80 my, consistent with the interpretation that Kyromyrma is a stem Formicinae. Другой муравей из Нью-джерсийского янтаря, Brownimecia clavata, был помещен первоначально в Ponerinae (Grimaldi et al. 1997) и позже в его собственном подсемействе в пределах УponeromorphsФ (Bolton 2003), a group roughly equivalent to the present
day УponeroidsФ plus the ectaheteromorphs. It seems probable that Brownimecia is also a crown group ant,
but uncertainty about relationships among the poneroids (see above) leaves open some room for doubt.
Конкурирующими кандидатами на звание самой древней группы кроны являются две ископаемые находки из Бирманского янтаря (Albian age, 100 mya): Burmomyrma rossi, возможно aneuretine (Dlussky 1996) и Myanmyrma gracilis, возможно myrmeciine (Engel & Grimaldi 2005; but see Archibald et al. 2006); и Gerontoformica cretacica (Nel et al. 2004) из французского янтаря сходного возраста. ќднако, Nel et al. (2004) трактует род Gerontoformica как incertae sedis
внутри Formicidae, но отмечает возможные сходства с Formicinae или Dolichoderinae. Вероятно, Burmomyrma и Gerontoformica или являются муравьями из кроны или очень близки к этой кладе. Они имеют габитус наводящий на размышления о современных муравьях, включая, в случае Gerontoformica, удлиненный скапус, который является характерным для муравьев группы кроны и отсутствующий у Sphecomyrma и Armaniines. ќстатки Burmomyrma найдены без головы, но другие черты их морфологии, и тот факт, что другие предположительно аневретины найдены в анадском янтаре (Campanian, 80 mya) (Engel & Grimaldi 2005), говорит в пользу рассмотрения его как группы кроны формицид.
Ископаемый род Myanmyrma имеет более озадачивающую специализированную абдоминальную морфологию (есть петиоль и постпетиоль), он имеет очень короткий scape, подобно другим муравьям стебля (stem ants).
Вероятно, группа кроны муравьиного эволюционного древа представляется младше по возрасту (Turonian, 90 mya) остатков из Ѕотсваны: Afropone и Afromyrma wбыли помещены в Ponerinae и Myrmicinae, соответственно (Dlussky et al. 2004), хотя это спорно (Wilson & Holldobler 2005,
Archibald et al. 2006).
Суммируя, на основании ископаемых находок весьма вероятно, что группа кроны (crown group) произошла примерно в позднем альбе (late Albian, ~100 mya) и они конечно присутствовали ранее турона (Turonian, 90 mya). There is considerable
diversity of body form among putative crown group ants from this period. Муравьи "стебля", или коренные (Stem ants, а конкретнее sphecomyrmines, armaniiines) также обнаруживаются в сходных по возрасту и тех же депозитах. Ранние находки этих групп известны из северного полушария (France, Myanmar), с более поздними появлениями в восточной северной Америке и южной Африке. Это доказывает, что муравьи произошли и диверсифицировались в Лавразии, до того как распространились в другие регионы (Perrichot et al. 2007).
It is worth belaboring the crown group/stem group distinction not only because it affects our thinking
about ant origins (and has been overlooked in previous discussions on this topic), but also because it becomes
important when incorporating fossil data into molecular divergence date estimates. When using a fossil to
constrain the minimum age of a node on a molecular tree the constraint applies to the stem group of the least
inclusive clade to which the fossil belongs (Magallуn 2004). Kyromyrma, for example, cannot be placed in
any subclade of Formicinae, so it supplies a minimum age estimate of 90 my for stem group Formicinae, i.e.,
for the node that represents the most recent common ancestor of this clade and its nearest extant relative. On
the other hand, the age estimates that are generated by divergence dating methods are typically reported as
crown group ages. Thus, an estimate for the age of crown group ants of, say, 115 my would not be contradicted
by the finding of stem ants (sphecomyrmines or armaniines) in older deposits.
Используя большой набор молекулярных данных по 7 ядерным генам 162 таксонов, Brady et al. (2006) установили параметры дивергенции основных групп муравьев by employing the penalized likelihood approach implemented in the program r8s (Sanderson 2003). Fossils were used to constrain the minimum ages of 41 nodes
within the tree, and the root node located deep within the aculeate wasp outgroups was fixed with two different
ages (one high, one low) reflecting a range of values consistent with the fossil record of Hymenoptera
as a whole. Установленный таким способом возраст для муравьев группы кроны (crown group ants) составляет около ~115 to 135 млн.лет, в полном соответствии с палеонтологической литературой (Grimaldi & Engel 2005), но противоречит предыдущим молекулярным исследованиям, которые вывели юрское происхождение муравьев (Crozier et al. 1997, Moreau, 2006).
Данные по дивергенции установленные для клад внутри Formicidae показали, что большинство подсемейств муравьев произошли (на уровне группы короны) в позднем мелу, followed by extensive within-subfamily diversification in
the Paleogene (Brady et al. 2006). Была выдвинута гипотеза, что муравьи расцвели и стали столь разнообразны вместе с появлением цветковых растений и что их эволюционный успех связан с обживанием комплексного лиственного слоя, образованного цветковыми - с доминированием лесов и симбиозом с сосущими растительные соки равнокрылыми насекомыми (Wilson & Holldobler 2005, Moreau, 2006). The fossil record and divergence date estimates are not inconsistent with this
hypothesis, но необходимы дополнительные исследования в этой области.
Высшая классификация
В первом томе "Системы природы" Карла Линнея (Systema Naturae. Linnaeus, 1758) описаны 17 видов муравьев (список здесь). Все они были помещены в состав одного рода Formica, хотя в течение нескольких десятилетий дополнительные роды были описаны или признаны, и эта тенденция продолжалась следующие годы, вместе с развитием более сложной иерархической классификации, в которой роды были распределены среди подсемейств и триб. поэтому, в итоге, муравьи описанные Линнеем (Linnaeus, 1758) сейчас распределены по 11 различным родам, принадлежащим к 4 подсемействам.
Всесторонний монументальный обзор всей истории изменений в высшей классификации муравьев было дано Болтоном (Bolton, 2003). Число подсемейств муравьев в большую часть 20 века изменялось от 7 до 10, с Aneuretinae, Cerapachyinae, Leptanillinae, Myrmeciinae и Pseudomyrmecinae, которые или признавались подсемействами, или входили в состав Dolichoderinae, Ponerinae, Dorylinae,
Ponerinae, Myrmicinae, соответственно. В последние 3 десятилетия наблюдается быстрое увеличение числа подсемейств как результат трех факторов: (1) некоторые подсемейства были искусственным объединением неродственных таксонов; (2) отказ от парафилетических таксонов, и (3) открытие новых ископаемых таксонов. В настоящее время признается 20 современных подсемейств и 5 ископаемых. Один ископаемый таксон (Armaniinae), часто имеет уровень отдельного семейства внутри надсемейства Formicoidea (Engel & Grimaldi 2005).
Таблица 1. Список валидных подсемейств и триб муравьев Formicidae (по: Engel & Grimaldi 2005 и Bolton et al. 2006). Подсемейства расположены в putative clades (Ward & Brady 2003, Brady et al. 2006), но размещение Leptanillinae, poneroids, и некоторых вымерших групп неясно. † = вымершие таксоны.
|
stem ants
|
†Armaniinae Dlussky (1983)
†Sphecomyrminae Wilson & Brown (1967)
†Haidomyrmecini Bolton (2003)
†Sphecomyrmini Wilson & Brown (1967) |
incertae sedis
|
Leptanillinae Emery (1910)
Anomalomyrmini Taylor (1990)
Leptanillini Emery (1910) |
poneroids
|
Agroecomyrmecinae Carpenter (1930)
Amblyoponinae Forel (1893)
†Brownimeciinae Bolton (2003)
Paraponerinae Emery (1901)
Ponerinae Lepeletier de Saint Fargeau (1835)
Platythyreini Emery (1901)
Ponerini Lepeletier de Saint Fargeau (1835)
Thaumatomyrmecini Emery (1901)
Proceratiinae Emery (1895)
Probolomyrmecini Perrault (2000)
Proceratiini Emery (1895)
|
formicoids: dorylomorphs
|
Aenictinae Emery (1901)
Aenictogitoninae Ashmead (1905)
Cerapachyinae Forel (1893)
Acanthostichini Emery (1901)
Cerapachyini Forel (1893)
Cylindromyrmecini Emery (1901)
Dorylinae Leach (1815)
Ecitoninae Forel (1893)
Cheliomyrmecini Wheeler, W. M. (1921)
Ecitonini Forel (1893)
Leptanilloidinae Bolton (1992)
|
formicoids: myrmeciomorphs
|
Myrmeciinae Emery (1877)
Myrmeciini Emery (1877)
Prionomyrmecini Wheeler, W. M. (1915)
Pseudomyrmecinae Smith, M. R. (1952)
|
formicoids: dolichoderomorphs
|
Aneuretinae Emery (1913)
Dolichoderinae Forel (1878)
Dolichoderini Forel (1878)
Iridomyrmecini Dubovikoff (2005) |
formicoids: ectaheteromorphs
|
Ectatomminae Emery (1895)
Ectatommini Emery (1895)
Typhlomyrmecini Emery (1911)
Heteroponerinae Bolton (2003) |
formicoids: Formicinae
|
Formicinae Latreille (1809)
Camponotini Forel (1878)
Dimorphomyrmecini Emery (1895)
Formicini Latreille (1809)
Gigantiopini Ashmead (1905)
Lasiini Ashmead (1905)
Melophorini Forel (1912)
Myrmecorhynchini Wheeler, W. M. (1917)
Myrmoteratini Emery (1895)
Notostigmatini Bolton (2003)
Oecophyllini Emery (1895)
Plagiolepidini Forel (1886) |
formicoids: Myrmicinae
|
Myrmicinae Lepeletier de Saint Fargeau (1835)
Adelomyrmecini Fernandez (2004)
Ankylomyrmini Bolton (2003)
Attini Smith, F. (1858)
Basicerotini Brown (1949)
Blepharidattini Wheeler & Wheeler (1991)
Cataulacini Emery (1895)
Cephalotini Smith, M. R. (1949)
Crematogastrini Forel (1893)
Dacetini Forel (1892)
Formicoxenini Forel (1893)
Lenomyrmecini Bolton (2003)
Liomyrmecini Bolton (2003)
Melissotarsini Emery (1901)
Meranoplini Emery (1914)
Metaponini Forel (1911)
Myrmecinini Ashmead (1905)
Myrmicini Lepeletier de Saint Fargeau (1835)
Paratopulini Bolton (2003)
Phalacromyrmecini Dlussky & Fedoseva (1988)
Pheidolini Emery (1877)
Solenopsidini Forel (1893)
Stegomyrmecini Wheeler, W. M. (1922)
Stenammini Ashmead (1905)
Tetramoriini Emery (1895) |
|
formicoids: others
|
†Formiciinae Lutz (1986)
†Paleosminthurinae Pierce & Gibron (1962) |
| |
Если же принять за главный принцип монофилетичность надвидовых таксонов, то это приведет к ситуации, намечающейся в группе кочевых муравьев дориломорф. Как известно, подсемейство Cerapachyinae, которое рассматривается как наиболее генерализованная группа дориломорфной клады, по молекулярно-генетическим оказывается немонофилетичным, что тперь имеет и морфологические обоснования (Brady & Ward 2005), и это уже очевидно (Brady 2003, Brady et al. 2006, Moreau et al. 2006). Все же, нет никакого простого решения этой проблемы, потому что точные детали церапахииновой парафилии остаются неясными. Есть недостаточное решение в основании дориломорфного древа, чтобы перестроить границы подсемейства, за исключением рассмотрения всей клады как одного подсемейства (Dorylinae).
Современные молекулярные исследования показали, что некоторые трибы внутри Formicinae (Lasiini, Plagiolepidini) и Myrmicinae (Dacetini, Pheidolini, Solenopsidini, Stenammini) немонофилетичны (Brady et al. 2006).
Классификация на родовом уровне более стабильна, ныне признаются 288 современных родов муравьев (Bolton et al. 2006). Большинство из них хорошо демаркированы и легко идентифицируются (Bolton
1994). Ќо остается ещЄ немало родов с очень широкой или слабой трактовкой, и которые явно парафилетичны, т.к. включают сателлитные группы.
Один из самых вопиющих случаев показывает род Pachycondyla (Ponerinae), который является почти синонимичным с целой трибой Ponerini. Интенсивный молекулярный анализ Chris Schmidt (pers. comm.), показал то, что и предполагалось: виды Pachycondyla разделяют и объединяют филогению всей трибы Ponerini, почти 20 родов. Это приведет к пересмотру классификации всей трибы. Однако, помещение всех видов в один род, объединяющий всю трибу Ponerini создаст огромный и сильно вариабельный род с примерно 1000 видами, что явно нежелательно. If we are to retain a Linnaean (i.e., ranked) phylogenetic classification system for ants, with named taxa that are recognizable with reasonable facility, then what is required hereЧand in other taxa where comparable problems ariseЧis a robust molecular phylogeny and identification of a set of mutually exclusive clades that are relatively easily
distinguished morphologically. This is a challenging task, and given considerable heterogeneity in rates of morphological evolution it remains unclear if this goal can always be achieved.
Многие крупные рода, рассматриваются как немонофилетичные: Amblyopone, Cerapachys,
Tetraponera, Camponotus, Trachymyrmex, Leptothorax, Nesomyrmex, Temnothorax, Aphaenogaster, Messor, Monomorium, Tetramorium. Resolving these into mutually exclusive monophyletic groups that can be
diagnosed with obvious phenotypic features will require comprehensive sampling of the relevant species,
selection of genes of appropriate variability, and careful scrutiny of morphological variation.
Таксономия на видовом уровне (Species-level taxonomy)
В первые годы было мало описаний новых таксонов муравьев, лишь работы первых мастеров: Linnaeus (1758), Fabricius, Latreille, Nylander, Westwood и др. Резкий рост начался после 1850 года, когда начались интенсивные исследования таких первых мирмекологов как F. Smith, Roger, и Mayr. Наибольшее количество новоописаний видов и подвидов муравьев приходится на начало 20 века, благодаря многочисленным таксономическим трудам классиков муравьиной систематики (Emery, Forel, Santschi, W. M. Wheeler).: все вместе они открыли почти треть современных видов - около 4000 таксонов, без учета синонимов]
Явный спад новоописаний набюдался в период 1930 по 1960. Надо отметить и такую особенность таксономии 19 и начала 20 веков: тогда выделяли множество инфравидовых форм и широкоочерченных видов. С появлением новой систематики в 1940-ых и 1950-ых, такие таксономисты как Creighton, Brown и Wilson стали учитывать большую внутривидовую изменчивость, отказываясь использовать подвиды (Buhs 2000, Ward 2007). По такому нетипологическому взгляду на вид, обнаружилось огромное количество синонимов, которые накопились в литературе и несмотря на продолжающееся существование в природе многих неописанных разновидностей в природе.
На сегодня в объеме мировой фауны ревизованы рода Neivamyrmex (Borgmeier 1955, Watkins
1976), Cataulacus (Bolton 1974a, 1982), Tetramorium (Bolton 1976, 1977, 1979, 1980), Rogeria (Kugler 1994), Gnamptogenys (Lattke 1995, 2004, Lattke et al. 2007), Cephalotes (de Andrade & Baroni Urbani 1999), Strumigenys (Bolton 2000), Pyramica (Bolton 2000), Proceratium (Baroni Urbani & de Andrade 2003) и Acropyga (LaPolla 2004). Конечно, и в них ещё продолжаются открытия. Но эти рода остаются наиболее ревизованными на видовом уровне по сравнению с другими крупными родами муравьев.
Такие крупнейшие рода как Crematogaster, Pheidole, Hypoponera, Paratrechina, Solenopsis,
и Camponotus требуют дальнейших и нелегких трудов для завершения их ревизий. Хотя некоторые из них имеют региональные ревизии (Trager 1984, Eguchi 2001, Longino 2003, Wilson 2003).
С мелкими родами ситуация много лучше, особенно в Европе (Seifert 2007) и Японии (Imai et al. 2003).
Появились онлайновые идентификаторы в интернете: AntWeb (www.antweb.org), Antbase (www.antbase.org), Australian Ants Online (www.ento.csiro.au/science/ants),
Ants of Costa Rica (http://academic.evergreen.edu/projects/ants/AntsofCostaRica.html) и Japanese Ant Image Database (http://ant.edb.miyakyo-u.ac.jp/E), Lucid (www.lucidcentral.org)... и мы начали создавать такой определитель]
Несколько современных технологических достижений обещают облегчить таксономию видового уровня. ќни включают усовершенствования в фотографировании и рисовании (например, Automontage system), измерения типовых экземпляров, картография распространения, и электронная организация данных. Расшифровка ДНК (DNA bar-coding) имеет потенциал, чтобы обеспечить ценную информацию о границах видов муравьев (Smith et al. 2005). Однако, технологический прогресс не упростит реальные природные взаимосвязи и не устранит трудности, возникающие у таксономиста и потребует всесторонних исследований. Недавнее обнаружение межвидовых социальных полиморфизмов (Krieger & Ross 2002), криптических разновидностей (Schlick-Steiner et al. 2006), взаимозависимых гибридных линий (Helms Cahan et al. 2002, Helms Cahan & Vinson 2003, Anderson et al. 2006), и клональная репродукция (Pearcy et al. 2004, Fournier et al. 2005) намекают на возможные осложнения, которые ждут новых иследователей таксономии муравьев, и выдвигают на первый план потребность и перспективность популяционных исследований при столкновении с таксономическими проблемами.
Благодарности
Работа поддержана University of California (Davis) и фондом US
National Science Foundation. Многие молекулярные выводы суммаризирована из Ant AToL (Assembling the Tree of Life) Project, поддержанного грантом NSF grant EF-0431330, при поддержки коллег Brian Fisher (California Academy of Sciences), Sean Brady, Ted
Schultz (Smithsonian Institution).

Реконструкция филогенеза муравьев по молекулярно-генетическим данным.
(©Science, по: Moreau et al., 2006)
–исунок в крупном виде
|
| |
ЛИТЕРАТУРА
Список литературы из оригинальной статьи. Отдельный список PDF статей здесь >>> а дополнительный список по молекулярной филогенетике см.ниже.
- Anderson, K.E., Gadau, J., Mott, B.M., Johnson, R.A., Altamirano, A., Strehl, C. & Fewell, J.H. (2006) Distribution and evolution of genetic caste determination in Pogonomyrmex seed-harvester ants. Ecology, 87, 2171-2184.
- Archibald, S.B., Cover, S.P. & Moreau, C.S. (2006) Bulldog ants of the Eocene Okanagan Highlands and history of the subfamily (Hymenoptera: Formicidae: Myrmeciinae). Annals of the Entomological Society of America, 99, 487-523.
- Baroni Urbani, C. & de Andrade, M.L. (2003) The ant genus Proceratium in the extant and fossil record (Hymenoptera: Formicidae). Museo Regionale di Scienze Naturali Monografie (Turin), 36, 1-492.
- Baroni Urbani, C., Bolton, B. & Ward, P.S. (1992) The internal phylogeny of ants (Hymenoptera: Formicidae).Systematic Entomology, 17, 301-329.
- Bergsten J. (2005) A review of long-branch attraction. Cladistics, 21, 163-193.
- Bolton, B. (1974) A revision of the Palaeotropical arboreal ant genus Cataulacus F. Smith (Hymenoptera: Formicidae). Bulletin of the British Museum (Natural History). Entomology, 30, 1-105.
- Bolton, B. (1976) The ant tribe Tetramoriini (Hymenoptera: Formicidae). Constituent genera, review of smaller genera and revision of Triglyphothrix Forel. Bulletin of the British Museum (Natural History). Entomology, 34, 281-379.
- Bolton, B. (1977) The ant tribe Tetramoriini (Hymenoptera: Formicidae). The genus Tetramorium Mayr in the Oriental and Indo-Australian regions, and in Australia. Bulletin of the British Museum (Natural History). Entomology, 36, 67-151.
- Bolton, B. (1979) The ant tribe Tetramoriini (Hymenoptera: Formicidae). The genus Tetramorium Mayr in the Malagasy region and in the New World. Bulletin of the British Museum (Natural History). Entomology, 38, 129-181.
- Bolton, B. (1980) The ant tribe Tetramoriini (Hymenoptera: Formicidae). The genus Tetramorium Mayr in the Ethiopian zoogeographical region. Bulletin of the British Museum (Natural History). Entomology, 40, 193-384.
- Bolton, B. (1982) Afrotropical species of the myrmicine ant genera Cardiocondyla, Leptothorax, Melissotarsus, Messor and Cataulacus (Formicidae). Bulletin of the British Museum (Natural History). Entomology, 45, 307-370.
- Bolton, B. (1990a) Abdominal characters and status of the cerapachyine ants (Hymenoptera, Formicidae). Journal of Natural History, 24, 53-68.
- Bolton, B. (1990b) The higher classification of the ant subfamily Leptanillinae (Hymenoptera: Formicidae). Systematic Entomology, 15, 267-282.
- Bolton, B. (1990c) Army ants reassessed: the phylogeny and classification of the doryline section (Hymenoptera, Formicidae). Journal of Natural History, 24, 1339-1364.
- Bolton, B. (1994) Identification guide to the ant genera of the world. Harvard University Press, Cambridge, Massachusetts, 222 pp.
- Bolton, B. (2000) The ant tribe Dacetini. Memoirs of the American Entomological Institute, 65, 1-1028.
- Bolton, B. (2003) Synopsis and classification of Formicidae. Memoirs of the American Entomological Institute, 71, 1-370.
- Bolton, B., Alpert, G., Ward, P.S. & Nasrecki, P. (2006) BoltonТs Catalogue of ants of the world. Harvard University Press, Cambridge, Massachusetts, CD-ROM.
- Borgmeier, T. (1955) Die Wanderameisen der neotropischen Region. Studia Entomologica, 3, 1-720.
- Brady, S.G. (2003) Evolution of the army ant syndrome: the origin and long-term evolutionary stasis of a complex of behavioral and reproductive adaptations. Proceedings of the National Academy of Sciences U. S. A., 100, 6575-6579.
- Brady, S.G., Fisher, B.L., Schultz, T.R. & Ward, P.S. (2006) Evaluating alternative hypotheses for the early evolution and diversification of ants. Proceedings of the National Academy of Sciences U. S. A., 103, 18172-18177.
- Brady, S.G. & Ward, P.S. (2005) Morphological phylogeny of army ants and other dorylomorphs (Hymenoptera: Formicidae). Systematic Entomology, 30, 593-618.
- Brown, W.L., Jr. (1954) Remarks on the internal phylogeny and subfamily classification of the family Formicidae. Insectes Sociaux, 1, 21-31.
- Brown, W.L., Jr. (1955) Ant taxonomy. In: Kessel, E.W. (Ed.) A century of progress in the natural sciences, 1853-1953. California Academy of Sciences, San Francisco, pp. 569-572.
- Brown, W.L., Jr., Gotwald, W.H., Jr. & Levieux, J. (1971) A new genus of ponerine ants from West Africa (Hymenoptera: Formicidae) with ecological notes. Psyche (Cambridge), 77, 259-275.
- Buhs, J.B. (2000) Building on bedrock: William Steel Creighton and the reformation of ant systematics, 1925-1970. Journal of the History of Biology, 33, 27-70.
- Carpenter, F.M. (1930) The fossil ants of North America. Bulletin of the Museum of Comparative Zoology, 70, 1-66.
- Carpenter, F.M. (1992) Treatise on invertebrate paleontology. Part R. Arthropoda 4. Volume 4. Superclass Hexapoda. Geological Society of America, Boulder, pp. i-ii, 279-655.
- Crozier, R.H., Jermiin, L.S. & Chiotis, M. (1997) Molecular evidence for a Jurassic origin of ants. Naturwissenschaften, 84, 22-23.
- de Andrade, M.L. & Baroni Urbani, C. (1999) Diversity and adaptation in the ant genus Cephalotes, past and present. Stuttgarter Beitrage zur Naturkunde. Serie B (Geologie und Paleontologie), 271, 1-889.
- de Queiroz, K. & Gauthier, J. (1990) Phylogeny as a central principle in taxonomy: phylogenetic definitions of taxon names. Systematic Zoology, 39, 307-322.
- Dlussky, G.M. (1999) The first find of the Formicoidea (Hymenoptera) in the lower Cretaceous of the northern hemisphere. [In Russian.] Paleontologicheskii Zhurnal, 1999(3), 62-66.
- Dlussky, G.M., Brothers, D.J. & Rasnitsyn, A.P. (2004) The first Late Cretaceous ants (Hymenoptera: Formicidae) from southern Africa, with comments on the origin of the Myrmicinae. Insect Systematics and Evolution, 35, 1-13.
- Dlussky, G.M. & Rasnitsyn, A.P. (2003) Ants (Hymenoptera: Formicidae) of Formation Green River and some other Middle Eocene deposits of North America. Russian Entomological Journal, 11, 411-436.
- Doyle, J.A. & Donoghue, M.J. (1993) Phylogenies and angiosperm diversification. Paleobiology, 19, 141-167.
- Eguchi, K. (2001) A revision of the Bornean species of the ant genus Pheidole (Insecta: Hymenoptera: Formicidae: Myrmicinae). Tropics Monograph Series, 2, 1-154.
- Emery, C. (1920) La distribuzione geografica attuale delle formiche. Tentativo di spiegarne la genesi col soccorso di ipotesi filogenetiche e paleogeografiche. Atti della Reale Accademia dei Lincei. Memorie. Classe di Scienze Fisiche, Matematiche e Naturali, (5)13, 357-450.
- Engel, M.S. & Grimaldi, D.A. (2005) Primitive new ants in Cretaceous amber from Myanmar, New Jersey, and Canada (Hymenoptera: Formicidae). American Museum Novitates, 3485, 1-23.
- Fournier, D., Estoup, A., Orivel, J., Foucaud, J., Jourdan, H., Le Breton, J. & Keller, L. (2005) Clonal reproduction by males and females in the little fire ant. Nature (London), 435, 1230-1234.
- Grimaldi, D. & Agosti, D. (2000a) The oldest ants are Cretaceous, not Eocene: comment. Canadian Entomologist, 132, 691-693.
- Grimaldi, D. & Agosti, D. (2000b) A formicine in New Jersey Cretaceous amber (Hymenoptera: Formicidae) and early evolution of the ants. Proceedings of the National Academy of Sciences U. S. A., 97, 13678-13683.
- Grimaldi, D., Agosti, D. & Carpenter, J.M. (1997) New and rediscovered primitive ants (Hymenoptera: Formicidae) in Cretaceous amber from New Jersey, and their phylogenetic relationships. American Museum Novitates, 3208, 1-43.
- Grimaldi, D. & Engel, M.S. (2005) Evolution of the insects. Cambridge University Press, Cambridge, xv + 755 pp.
- Helms Cahan, S., Parker, J.D., Rissing, S.W., Johnson, R.A., Polony, T.S., Weiser, M.D. & Smith, D.R. (2002) Extreme genetic differences between queens and workers in hybridizing Pogonomyrmex harvester ants. Proceedings of the Royal Entomological Society of London. Series B, 269, 1871-1877.
- Helms Cahan S. & Vinson S.B. (2003) Reproductive division of labor between hybrid and non hybrid offspring in a fire ant hybrid zone. Evolution, 57, 1562-1570.
- Holldobler, B. & Wilson, E.O. (1990) The ants. Harvard University Press, Cambridge, Massachusetts, xii + 732 pp.
- Imai, H. T., Kihara, A., Kondoh, M., Kubota, M., Kuribayashi, S., Ogata, K., Onoyama, K., Taylor, R. W., Terayama, M., Tsukii, Y., Yoshimura, M. & Ugawa, Y. (2003) Ants of Japan. Gakken, Tokyo, 224 pp.
- Krieger, M.J. & Ross, K.G. (2002). Identification of a major gene regulating complex social behavior. Science (Washington, D.C.), 295, 328-332.
- Kugler, C. (1994) A revision of the ant genus Rogeria with description of the sting apparatus (Hymenoptera: Formicidae). Journal of Hymenoptera Research, 3, 17-89.
- LaPolla, J.S. (2004) Acropyga (Hymenoptera: Formicidae) of the world. Contributions of the American Entomological Institute, 33(3), 1-130.
- Lattke, J.E. (1995) Revision of the ant genus Gnamptogenys in the New World (Hymenoptera: Formicidae). Journal of Hymenoptera Research, 4, 137-193.
- Lattke, J.E. (2004) A taxonomic revision and phylogenetic analysis of the ant genus Gnamptogenys Roger in Southeast Asia and Australasia (Hymenoptera: Formicidae: Ponerinae). University of California Publications in Entomology, 122, 1-266.
- Lattke, J.E., Fernandez, F. & Palacio, E.E. (2007). Identification of the species of Gnamptogenys Roger in the Americas. Memoirs of the American Entomological Institute, 80, 254-270.
- Linnaeus, C. (1758) Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I. Editio decima, reformata. L. Salvii, Holmiae [= Stockholm], 824 pp.
- Longino, J.T. (2003) The Crematogaster (Hymenoptera, Formicidae, Myrmicinae) of Costa Rica. Zootaxa, 151, 1-150.
- Magallуn, S. (2004) Dating lineages: molecular and paleontological approaches to the temporal framework of clades. International Journal of Plant Science, 165(4 Suppl.), S7-S21.
- Masuko, K. (1990) Behavior and ecology of the enigmatic ant Leptanilla japonica Baroni Urbani (Hymenoptera: Formicidae: Leptanillinae). Insectes Sociaux, 37, 31-57.
- Moreau, C.S., Bell, C.D., Vila, R., Archibald, S.B. & Pierce. N.E. (2006) Phylogeny of the ants: diversification in the age of angiosperms. Science (Washington D. C.), 312, 101-104.
- Morley, B.D.W. (1938) An outline of the phylogeny of the Formicidae. Bulletin de la Societe Entomologique de France, 43, 190-194.
- Ouellette, G.D., Fisher, B.L. & Girman, D.J. (2006) Molecular systematics of basal subfamilies of ants using 28S rRNA(Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution, 40, 359-369.
- Pearcy, M., Aron, S., Doums, C. & Keller, L. (2004) Conditional use of sex and parthenogenesis for worker and queen production in ants. Science (Washington D. C.), 306, 1780-1783.
- Perrichot, V., Lacau, S., Nuraudeau, D. & Nel, A. (2007) Fossil evidence for the early ant evolution. Naturwissenschaften.
- Poinar, G., Jr., Archibald, B. & Brown, A. (1999) New amber deposit provides evidence of early Paleogene extinctions, paleoclimates, and past distributions. Canadian Entomologist, 131, 171-177.
- Poinar, G., Baroni Urbani, C. & Brown, A. (2000) The oldest ants are Cretaceous, not Eocene: reply. Canadian Entomologist, 132, 695-696.
- Sanderson M.J. (2003) r8s: inferring absolute rates of molecular evolution and divergence times in the absence of a molecular clock. Bioinformatics, 19, 301-302.
- Saux, C., Fisher, B.L. & Spicer, G.S. (2004) Dracula ant phylogeny as inferred by nuclear 28S rDNA sequences and implications for ant systematics (Hymenoptera: Formicidae: Amblyoponinae). Molecular Phylogenetics and Evolution, 33, 457-468.
- Seifert, B. (2007) Die Ameisen Mittel- und Nordeuropas. lutra Verlags- und Vertriebsgesellschaft, Tauer, 368 pp.
- Schlick-Steiner, B.C., Steiner, F.M., Moder, K., Seifert, B., Sanetra, M, Dyreson, E., Stauffer, C. & Christian, E. (2006) A multidisciplinary approach reveals cryptic diversity in western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Molecular Phylogenetics and Evolution, 40, 259-273.
- Smith, M.A., Fisher, B.L. & Hebert, P.D.N. (2005) DNA barcoding for effective biodiversity assessment of a hyperdiverse arthropod group: the ants of Madagascar. Philosophical Transactions of the Royal Society of London. B. Biological Sciences, 360, 1825-1834.
- Taylor, R.W. (1978) Nothomyrmecia macrops: a living-fossil ant rediscovered. Science (Washington D. C.), 201, 979-985.
- Trager, J.C. (1984) A revision of the genus Paratrechina (Hymenoptera: Formicidae) of the continental United States. Sociobiology, 9, 49-162.
- Ward, P.S. (1994) Adetomyrma, an enigmatic new ant genus from Madagascar (Hymenoptera: Formicidae), and its implications for ant phylogeny. Systematic Entomology, 19, 159-175.
- Ward, P.S. (2007) Edward O. Wilson and his contributions to ant systematics. Memoirs of the American Entomological Institute, 80, 3-7.
- Ward, P.S. & Brady, S.G. (2003) Phylogeny and biogeography of the ant subfamily Myrmeciinae (Hymenoptera: Formicidae). Invertebrate Systematics, 17, 361-386.
- Ward, P.S. & Downie, D.A. (2005) The ant subfamily Pseudomyrmecinae (Hymenoptera: Formicidae): phylogeny and evolution of big-eyed arboreal ants. Systematic Entomology, 30, 310-335.
- Watkins, J.F., II. (1976) The identification and distribution of New World army ants (Dorylinae: Formicidae). Baylor University Press, Waco, Texas, 102 pp.
- Wheeler, W.M. (1920) The subfamilies of Formicidae, and other taxonomic notes. Psyche (Cambridge), 27, 46-55.
- Wilson, E.O. (2003) Pheidole in the New World. A dominant, hyperdiverse ant genus. Harvard University Press, Cambridge, Massachusetts, [ix] + 794 pp.
- Wilson, E.O., Carpenter, F.M. & Brown, W.L., Jr. (1967) The first Mesozoic ants, with the description of a new subfamily. Psyche (Cambridge), 74, 1-19.
- Wilson, E.O. & Holldobler, B. (2005) The rise of the ants: a phylogenetic and ecological explanation. Proceedings of the National Academy of Sciences U. S. A., 102, 7411-7414.
Список литературы по молекулярно-филогенетической теме.
-
Baker, R. H., Wilkinson, G. S., and DeSalle, R (2001) Phylogenetic utility of different types of molecular data used to infer evolutionary relationships among stalk-eyed flies (Diptera). Syst. Biol. 50, 87-105.
-
Baroni Urbani, C, Bolton, B., and Ward, P. S. (1992) The internal phylogeny of ants (Hymenoptera: Formicidae). Syst. Entomol. 17, 301-329.
-
Bolton, B. (1994) Identification Guide to tlie Ant Genera of the World. Harvard University Press: Cambridge, MA, USA.
-
Brown, W. L J. (1954) Remarks on the internal phylogeny and subfamily classification of the family Formicidae. Insectes Sociaux 1, 21-31.
-
Dowton, M., and Austin, A. D. (1994) Molecular phylogeny of the insect order Hymenoptera: Apocritan relationships. Proc. Natl. Acad. Sci. USA 91, 9911-9915.
-
Dowton, M., and Austin, A. D., (2002) Increased congruence does not necessarily indicate increased phylogenetic accuracy -the behavior of the incongruence length difference test in mixed-model analyses. Syst. Biol. 51, 19-31.
-
Farris, J. S., Kallersjo, M., Huge, A. G., and Bult, — (1995) Testing significance of incongruence. Cladistics 10, 315-319
-
Felsenstein, J. (1981) Evolutionary trees from DNA sequences: a maximum likelihood approach. J. Mol. Evol. 17, 368-376.
-
Felsenstein, J. (1985) Confidence limits on phylogenies: an approach using bootstrap. Evolution 39, 783-791.
-
Fitch, W. M. (1971) Toward defining the course of evolution: minimum change for a specified tree topology. Syst. Zool. 20, 406-416.
-
Gimeno C, Belshaw, R., and Quicke D. L. (1997) Phylogenetic relationships of the Alysiinae/Opiinae (Hymenoptera: Brac-onidae) and the utility of cytochrome b, 16S and 28S D2 rRNA. Insect Mol. Biol. 6, 273-284.
-
Hassouna, N.. Michot B., and Bachellerie, J. P. (1984) The complete nucleotide sequence of mouse 28S rRNA gene. Implications for the process of size increase of the large subunit rRNA in higher eukaryotes. Nucleic Acids Res 12, 3563-3583.
-
Holldobler, B., and Wilson, E. ќ. (1990) The Ants. The Belknap Press of Harvard University Press, Cambridge, Mass.
-
Kimura, M. (1980) A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 16,111-120.
-
Long, E. O., and Dawid, I. B. (1980) Repeated genes in eukaryotes. Annu. Rev. Biochem. 49, 727-726.
-
Ohnishi, H., and Yamamoto. M-T. The structure of a single unit of ribosomal RNA gene (rDNA) including intergenic subrepeats in the Australian bulldog ant Myrmecia croslandi (Hymenoptera: Formicidae). Zool. Sci. in press.
-
Pelandakis, M., and Solignac, M. (1993) Molecular phylogeny of Drosophilc based on ribosomal RNA sequences. J. Mol. Evol. 37, 525-543.
-
Posada, D., and Crandell, K. A. (1998) Modeltest: testing the model of DNA substitution. Bioinform. 14, 817-818.
-
Posada, D., and Crandell, K. A. (2001) Selecting the best-fit model of nucleotide substitution. Syst. Biol. 50, 580-601.
-
Rodriguez, F., Oliver, J. L., Marin, A., and Medina, J. R. (1990) The general stochastic model of nucleotide substitution. J. Theor. Biol. 142, 485-501.
-
Saitou, N., and Nei, M. (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406-425.
-
Sambrook, J., Fritsch, E . F., and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
Schmitz, J., and Moritz, R. F. A. (1998) Molecular phylogeny of Vespidae (Hymenoptera) and the evolution of sociality in wasps. Mol. Phylogenet Evol. 9, 183-191.
-
Sullender, B.W. (1998) A molecular phylogeny for the Formicidae. http://www. ruf.rice.edu/-bws/molecular_phylogeny.html
-
Swofford, D. L. (2002) PAUP*. Phylogenetic analysis using parsimony (*and other methods), version 4.0. Sinauer associates: Sunderland, MA, USA.
-
Tamura, K., and Nei, M. (1993) Estimation of the number of nucleotide substitutions in control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol Evol 10, 512-526.
-
Taylor. R. W. (1978) Nottiomyrmecia mccrops: a living-fossil ant rediscovered. Science 201, 979-985.
-
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673-4680.
-
Ward. P. S., and Brady, S. G. (2003) Phylogeny and biogeography of the ant subfamily Myrmeciinae (Hymenoptera: Formicidae). Invertebrate Systematics 17, 361-386.
-
Wilson, E. 0. (1971) The Insect Societies. Harvard University Press: Cambridge, MA, USA.
-
Yoder, A. D., Irwin, J. A., and Payseur, B. A. (2001) Failure of the ILD to determine data combinability for slow loris phylogeny. Syst. Entomol. 50, 408-424.
|
| |
| |
|
©2008, Vladislav Krasilnikov (translation & supplement)
|
|
Всякое использование без согласования с автором и без активной гиперссылки на наш сайт преследуетс¤ в соответствии с Российским законодательством об охране авторских прав.
|
