
Муравьиные новости (2004 - №40)

***
"Insectes sociaux", Муравьиные
НОВОСТИ: Виды-2022 и 2004,
Статьи-2005 и 2004
|
Добавлено 29-5-2005
|
|
Formica picea: отдельный от Formica candida вид.
|
|
Bernhard SEIFERT
E-mail:bernhard.seifert@smng.smwk.sachsen.de
Staatliches Museum fur Naturkunde Gorlitz, Postfach 300154, D-02806 Gorlitz,
Germany. E-mail:
The "Black Bog Ant" Formica picea NYLANDER, 1846 – a species different from
Formica candida SMITH, 1878 (Hymenoptera: Formicidae).
|
|
"Myrmecologische Nachrichten",
("Myrmecological News")
2004, Volume 6, pp.29 - 38, Wien, Dezember 2004.
|
|
|
Ревизована таксономия черного болотного муравья "Black Bog Ant". Согласно морфометрическим и генетическим данным, таксон собранный BOLTON (1995) под
именем Formica candida SMITH, 1878 должен быть разделен как минимум на 2 различных вида, отличающихся ещё и зоогеографией. Болотный муравей,
распространенный в Европе, на Кавказе и в Западной Сибири идентифицирован как Formica picea NYLANDER, 1846, а вид, распространенный в горах Центральной
Азии на север до Горно-Алтайска, в Тибете, Монголии, Прибайкалье, Восточной Сибири - переописывается как Formica candida SMITH, 1878 с фиксацией
неотипа. Согласно статье 23.9.5. 4-го издания Международного Кодекса Зоологической Номенклатуры ICZN (1999), название Formica picea NYLANDER, 1846 (a
junior primary homonym of F. picea LEACH, 1825) не нуждается в замене ни одним из его младших синонимов. Это правило более применимо, так как
фактически F. picea LEACH находится в составе рода Camponotus MAYR с 1861 года и никогда после этого не возвращался обратно в род Formica; это положит
конец, длящейся почти 50 лет таксономической путанице между тремя именами. Приводятся таблицы с подробными морфометрическими данными по разделению всех
Палеарктических черных блестящих с редуцированной опушенностью муравьев рода Formica: F. picea, F. candida, F. gagatoides RUZSKY, 1904, F. kozlovi
DLUSSKY, 1965 и F. gagates LATREILLE, 1798.
Таблица сравнения признаков F. candida и F. picea
Подробности о роде Formica читайте ЗДЕСЬ >>>, а о подсемействе Formicinae ЗДЕСЬ >>>.
***
|
|
|
| |
|
ВВЕДЕНИЕ
Проведено исследование сравнительного коллекционного материала по одному из известных в Европе и Сибири видов муравьев. Черные блестящие виды подрода
Serviformica, известные мирмекологам как черные болотные муравьи "Black Bog Ant", это один из худших примеров непостоянного и неопределенного
таксономического поименования в этой группе. В Европе, но не за её пределами, этот муравей один из немногих реальных болотных tyrphophiles среди
муравьев, хотя в высокогорьях он селится и на плотных почвах (e.g., KUTTER 1917). Его специфические условия обитания и блестящее тело создали редкую
ситуацию, что большинство европейских находок по этому муравью, опубликованные за последние 150 лет, относятся к одному и тому же виду. Как следствие,
мы имеем довольно хорошие знания его биологии.
С 1846 по 1979 годы эти муравьи почти всегда именовались как Formica picea NYLANDER, 1846. Учитывая младшую первичную гомонимию F. picea
NYLANDER, 1846 с Formica picea LEACH, 1825 (с 1861 в составе Camponotus MAYR), во 2-й половине XX века были предложены различные младшие синонимы как
замещающие имена.
Сначала YARROW (1954) первым предожил замещающее название Formica transkaucasica NASSONOV, 1889. Этому предложению не следовали большинство
мирмекологов - очевидно, потому что они не рассматривали гомонимию как практическую проблему (Dlussky 1967, Dlussky & Pisarski 1971, Kutter 1977,
Arnoldi & Dlussky
1978). Имя F. transkaucasica стало чаще использоваться тогда, когда COLLINGWOOD (1979) повторил предложенное Yarrow имя в его широко распространенной
книге по муравьям Фенноскандии.
Наконец, последнее измение, предложенное BOLTON (1995), ввело в обиход название F. candida SMITH, 1878 после выхода его мирового каталога всех
видов муравьев.
Но все эти изменения, предложенные тремя уважаемыми английскими мирмекологами (Yarrow, Collingwood, Bolton) не были основаны (!) ни на
исследовании типового материала, ни на критическом прочтении оригинальных описаний, ни на изучении сходных видов в типовых местностях. Теоретическая
астрономия чистой воды.
Следовательно, мы столкнулись с проблемой, что никто сейчас не знает какое оригинальное описание (или F. transkaucasica, или F. candida) должно
применяться к реальному виду черных болотных муравьев Black Bog Ant.
Согласно и генетическим свидетельствам (GOROPASHNAYA, 2003) и морфологическим данным, представленным здесь, Палеарктичесий комплекс всех таксонов,
отнесенных BOLTON (1995) к виду Formica candida SMITH, 1878, состоит по-крайней мере, из 2-х отдельных видов. Переописание и фиксация неотипа
Центрально-Азиатско-Тибетского вида Formica candida и переописание и фиксация лектотипа Европейско-Западно-Сибирского Черного болотного муравья Formica
picea NYLANDER, 1846, предствлено как первый шаг в наведении таксономического порядка в этом комплексе видов.
МАТЕРИАЛ
Автор изучил 264 рабочих муравья из 105 гнезд (материал хранится в Museum fur Naturkunde Gorlitz) 5 видов
Formica picea NYLANDER, 1846, F. candida SMITH, 1878, F. gagatoides RUZSKY, 1904, F. kozlovi DLUSSKY, 1965, и F. gagates LATREILLE, 1798.
All measurements were made on mounted and
dried specimens using a goniometer-type pin-holding
device, permitting endless rotations around X, Y,
and Z axes. A Wild M10 stereomicroscope equipped
with a 1.6 x planapochromatic objective was used at
magnifications of 50 - 320 x. A mean measuring error
of +/- 1 µm is given for small and well-defined structures,
such as hair length, but may reach 4 µm for
measures > 1700 µm with difficult positioning and
high influence of air humidity. To avoid rounding errors,
all measurements were recorded in µm even
for characters for which a precision of +/- 1 µm is impossible.
Error sources of stereomicroscopic measuring
in general and of the particular system used here
are properly discussed elsewhere (SEIFERT 2002).
Setae, also called pilosity or simply "hairs", are
differentiated from pubescence hairs in having a
much larger diameter – usually 4 - 8 мm in setae
and 1 - 2 µm in pubescence. All seta counts (acronyms
beginning with "n") are restricted to standing
setae projecting > 10 µm from cuticular surface as
seen in a profile view specifically defined.
ПРОМЕРЫ
Определение числовых признаков и индексов:
CL maximum cephalic length in median line; the
head must be carefully tilted to the position
with the true maximum. Excavations of occiput
and/or clypeus reduce CL.
CS cephalic size; the arithmetic mean of CL and
CW, used as a less variable indicator of body
size.
CW maximum cephalic width; this is either across,
behind, or before the eyes.
EYE eye-size: the arithmetic mean of the large
(EL) and small diameter (EW) of the elliptic
compound eye.
GHL length of longest seta on dorsal plane of first
gaster tergite excluding the row of setae immediately
anterior of the hind tergite margin.
nGU unilateral number of setae protruding more
than 10 µm from cuticular surface of the underside
of head ("gula") as visible in lateral view.
nHFFL arithmetic mean of the number of setae protruding
more than 10 µm from cuticular surface
of the flexor profile of hind femora.
nMN unilateral number of setae on mesonotum protruding
more than 10 µm from cuticular surface.
nOCC with the head in measuring position for CL,
unilateral number of setae protruding more
than 10 µm from occipital margin and the head
sides anterior to level of anterior eye margin.
nPE unilateral number of setae protruding more
than 10 µm from margin of petiole scale dorsal
of spiracle in caudal or frontal viewing
position.
nPN unilateral number of setae protruding more
than 10 µm from cuticular surface of pronotum.
nPR unilateral number of setae protruding more
than 10 µm from cuticular surface on propodeum
and lateral metapleuron (excluding setae
fringing the metapleural gland orifice and
those on ventrolateral edge of metapleuron).
PEW maximum width of petiole.
RipD average distance of transversal microripples
on dorsal plane of first gaster tergite. At least
3 countings along a 90 мm distance on different
surface spots are averaged – in species
of the F. candida complex 5 - 8 coun
objectives with numeric apertures > 0.200,
clean surfaces and light inclined perpendicular
to the ripples. Do not include anastomosae
between the ripples into the counting.
SL maximum straight line scape length excluding
the articular condyle.
sqPDG square root of pubescence distance on dorsum
of first gaster tergite. The number of
pubescence hairs n crossing a transverse
measuring line of length L is counted, hairs
just touching the line are counted as 0.5.
The pubescence distance PDG is then given
by L / n. In order to normalise positively
skewed distributions, the square root of PDG
is calculated. Exact counting is promoted by
clean surfaces and flat, reflexion-reduced il
lumination directed perpendicular to the axis
of pubescence hairs. Use high-resolution objectives
with numeric apertures > 0.200. In
each specimen 4 - 6 measuring-lines of 400 мm
are averaged under exclusion of surface parts
with apparently detached pubescence. If there
are no transects with undamaged pubescence
possible, PDG can be calculated by the formula
PDG = BD2 / PLG where BD is the
mean distance of hair base punctures and PLG
the mean length of pubescence hairs.
In most of the species groups of Formica, body
ratios are strongly influenced by allometric growth. In
order to make numeric characters directly comparable
between the species, predictions for the assumption
of all individuals having an identical cephalic size
of 1.4 mm were calculated with the following equations
calculated as mean of 24 Palaearctic species of
the subgenus Serviformica:
CL / CW1.4 = CL / CW / (-0.1143 * CS + 1.2936) * 1.1336
SL / CS1.4 = SL / CS / (-0.1077 * CS + 1.2062) * 1.0554
EYE / CS1.4 = EYE / CS / (-0.0594 * CS + 0.3752) * 0.292
GHL / CS1.4 = GHL / CS / (-0.0008 * CS + 0.0852) * 0.0841
PEW / CS1.4 = PEW / CS / (0.1001 * CS + 0.3039) * 0.444
nOCC1.4 = nOCC / (3.268 * CS + 1.31) * 5.88
nGU1.4 = nGU / (1.635 * CS + 0.43) * 2.72
nPN1.4 = nPN / (10.81 * CS - 6.50) * 8.63
nMN1.4 = nMN / (6.10 * CS - 5.21) * 3.33
nPR1.4 = nPR / (5.50 * CS - 3.97) * 3.73
nPE1.4 = nPE / (3.94 * CS - 1.80) * 3.71
nHFFL1.4 = nHFFL / (4.13 * CS - 0.45) * 5.34
sqPDG1.4 = sqPDG / (0.953 * CS + 3.086) * 4.42
RipD1.4 = RipD / (-0.0632 * CS + 7.29) * 7.2
Negative or positive signs in the divisor indicate
a negative or positive allometry of the ratios but not
necessarily in the other characters.
Таксономия
Formica picea NYLANDER, 1846
Исследованный материал: Австрия, Германия, Грузия, Чехия, Франция, Швеция, Россия (Звенигород и Баксан на Северном Кавказе), всего 62 муравья
из 26 сборов
ОПИСАНИЕ
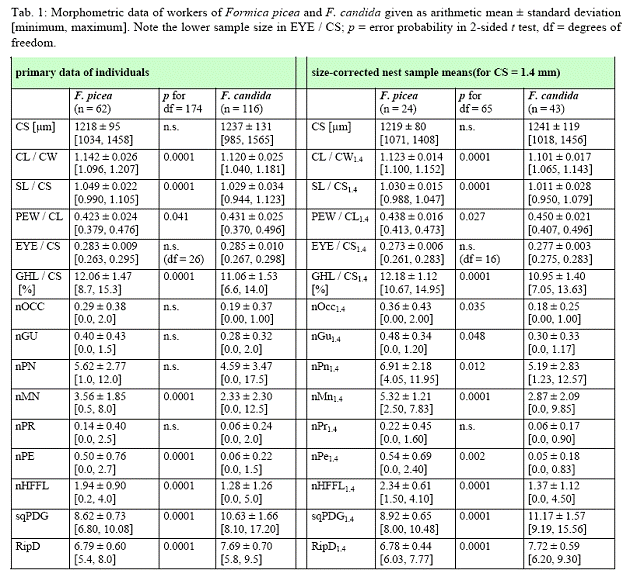
Описание рабочих особей по Зейферту (B.SEIFERT, 2004): "Mean size distinctly
smaller than Serviformica average: CS 1.22 mm. Head
and scape significantly longer than in F. candida:
CL / CW1.4 1.123, SL / CS1.4 1.030 (Tab. 1). Frontal
triangle as shining as the adjacent surfaces. Cuticular
surface of head, mesosoma and gaster as result of
reduced pubescence and weak microsculpture shining;
a denser pubescence is usually developed only on
propodeum and petiole. However, pubescence and
microsculpture significantly denser than in F. candida:
on dorsal surface of first gaster tergite, average distance
of transverse microripples 6.8 мm (RipD) and
of pubescence hairs 74 мm (PDG). Clypeus, procoxae,
ventral parts of meso- and metacoxae and all
ventral and dorsal gaster sclerites with quite numerous
and very long erect setae which are longer than
in F. candida, GHL / CS 12.1 %. Setae numbers a
little larger than in F. candida: Seen in lateral profile,
hind vertex with 3 - 16 and central vertex with
2 - 7 setae. Both sides of pronotum usually with 5 -
17 and of mesonotum with 3 - 11 long, erect setae.
Propodeum occasionally and petiole scale usually
with single short setae. Flexor sides of both hind tibiae
with a sum of 2 - 6 semierect to subdecumbent
setae. Petiole scale variable, on the average narrower
than in F. candida; its dorsal margin in frontal view
fully convex or with a straight or concave median part;
scale in lateral view not very thick with a tapering
apex. Colouration pattern similar to situation in F.
candida. For morphometric details of 62 worker specimens
see Tab. 1." (B.SEIFERT, 2004)
Таксономия
Formica transkaucasica NASSONOV, 1889 –
как Formica incertae sedis
Типовые экземпляры не найдены, видимо утеряны (в статье приводится ссылка на бывшего и нынешнего кураторов Зоологического Музея МГУ - Длусского
и Антропова). Так что, с этим названием (как неопределенным) мы снова и, видимо, надолго расстаемся.
Таксономия
Formica candida SMITH, 1878
Исследованный материал: Бутан, Индия (Кашмир), Казахстан, Киргизия, Китай, Монголия, Россия (Алтай, Прибайкалье, район Уссури, Якутия), всего
117 муравьев из 44 сборов
ОПИСАНИЕ
Описание рабоих особей по Зейферту (B.SEIFERT, 2004): "Mean size smaller than
Serviformica average: CS 1.240 mm. Head and scape
shorter than in the next related species, i.e. F. picea,
F. gagatoides and F. kozlovi: CL / CW1.4 1.101, SL
/ CS1.4 1.010 (Tab. 2). Frontal triangle as shining as
the adjacent surfaces. Cuticular surface of head, mesosoma
and gaster, as result of strongly reduced pubes36
cence and very weak microsculpture, very shining;
a denser pubescence is usually developed only on propodeum
and petiole. On dorsal surface of first gaster
tergite, average distance of transverse microripples
7.7 мm (RipD) and of pubescence hairs 113 мm
(PDG). Clypeus, procoxae, ventral parts of meso- and
metacoxae and all ventral and dorsal gaster sclerites
with quite numerous and very long erect setae. Seen
in lateral profile, hind vertex with 3 - 7 and central
vertex with 2 - 8 setae. Both sides of pronotum usually
with 2 - 16 and of mesonotum with 1 - 9 long,
erect setae. Propodeum and petiole scale only very
occasionally with single short and weak setae. Flexor
sides of both hind tibiae with a sum of 0 - 5 semierect
to subdecumbent setae. Petiole scale variable, its dorsal
margin in frontal view fully convex or with a
straight or concave median part; scale in lateral view
not very thick and with a tapering apex. An average
colouration pattern is: head, mesosoma, coxae and
gaster blackish brown; appendages lighter, mandibles
light to dark reddish brown. Slightly lighter or darker
specimens may occur. For morphometric details of
117 worker specimens see Tab. 1." (B.SEIFERT, 2004)
Formica candida был описан по одной единственной самке найденной "... On the road across
the Pamir, from Sarikol to Panja ..." (SMITH 1878). По информации кураторов коллекций Университетского Музея в Оксфорде (Oxford University Museum) и
Британском Музее (British Museum of Natural
History) местонахождение этого типового материала неизвестно (Chris O'Toole and Barry Bolton, pers.
comm., 2001). Запросы, посланные Национальной Зоологической Службе Индии в Калькутте (National Zoological Survey of India Calcutta), другого возможного
хранилища, ответа не имели.
Поэтому сам автор новой статьи произвел фиксацию НЕОТИПА:
Экземпляр этикетирован "KYRGHYZTAN: Alai-Valley 39.42.09 N, 73.27.24 E, 3200 m, leg. R.Schultz
1999.07.21-67" и "Neotype Formica candida Smith 1878 det. Seifert". Это нижний экземпляр муравьев из трех рабочих особей на той же булавке, собранные
из одного гнезда и сохраняемые в музее SMN Gorlitz.
Неотип имеет промеры CS 1.112 mm, CL / CW 1.132, SL
/ CS 1.032. Неотип найден на расстоянии около 315 km N от места где предположительно был найден тип Смсита (Smith 1878), как "very near". Данные по ДНК
(mtDNA sequence data) неотипа No. 67 сохранены в Gen-Bank Accession No. AY786154 (Haplotype "Kyrgizstan-II"), также как и экземпляры No. 64 и No. 66
из места находки неотипа под номером No. AY786153 (Haplotype "Kyrgizstan-I").
Таксономия
ОТЛИЧИЯ ВИДОВ
Читайте через неделю другие подробности
Согласно данным по исследованию трех локусов митохондриальных ДНК (mtDNA loci) с 1500 пар оснований (GOROPASHNAYA, 2003), комплекс видов, собранный
Болтоном (BOLTON, 1995) под именем F. candida complex показывает явную филогеографическую структуру и может быть разделен на две основные клады –
Европейско-Западно-Сибирскую кладу и кладу гор Центральной Азии.
Эти 2 клады демонстрируют дивергенцию (net sequence divergence) в 1.2 %, что даже больше, чем это известно в роде Formica. Даже, такие хорошо отличимые
виды как F. pratensis и F. lugubris, или F. truncorum и F. frontalis, отличаются только в пределах от 0.85 до 1.2 % (GOROPASHNAYA & al. 2004a,
b).
Генетические отличия Formica picea и Formica candida подтверждаются на морфологическом уровне дискриминантным анализом используя абсолютный размер
головы (cephalic size) и 13 признаков скорректированных с размером по экземплярам из 67 гнезд (как минимум 2 рабочих на выборку).
Дискриминант D(14):
D(14) = 42.98 - 2.65 CS - 44.11 CL / CW1.4 +
0.58 SL/CS1.4 - 8.88 PEW / CS1.4 + 4.18 GHL /
CS1.4 + 0.12 nOCC1.4 + 0.79 nGU1.4 + 0.48 nPN1.4
- 0.54 nMN1.4 + 0.29 nPR1.4 - 1.25 nPE1.4 - 0.25
nHFFL1.4 + 0.72 RipD1.4 + 0.67 sqPDG1.4
предполагает следующее разделение Палеарктических популяций на 2 группы, которые здесь интерпретируются как отдельные виды Formica picea и Formica
candida.
F. picea: D(14) -2.049 ± 0.894 [-0.49,-3.58] n = 24
F. candida: D(14) 2.054 ± 1.053 [0.49, 4.81] n = 43
Причем, автор заявляет, что кластер F. candida может быть в дальнейшем разделен ещё на двое и тогда понадобтся вернуть и третье название (Рузского,
Фореля, Штарка, Насонова?).
Отличия по дискриминанту
Вид Formica gagates может быть отличен от близких видов комбинацией признаков из экстремально крупных SL / CS, GHL / CS, менее крупного sqPDG и
крупного RipD. Дифференциации помогает и их экология и зоогеография. Большой кластер из видов F. gagatoides + F. kozlovi и F. candida + F. picea имеет
более низое GHL/ CS, sqPDG ималое число щетинок на промезонотуме у первых двух видов. Типы части видов хранятся в России (лектотип F. gagatoides в
Зооологическом музее ЗИН РАН, Санкт-Петербург, голотип F. kozlovi - в Зоологическом музее МГУ, Москва). Вид Formica kozlovi отличается от F.
gagatoides значительно более длинным скапусом, более крупным nPN и меньшим RipD. Вид F. kozlovi также отличается степенью опушенности на 1-м и 2-м
тергите брюшка (у F. gagatoides она сходная на двух этих тергитах).
Таблица сравнения признаков F. candida и F. picea (SEIFERT, 2004)
***
|
|
©2005, Владислав Красильников (перевод, комментарии). Всякое использование без согласования с автором и без активной
гиперссылки на наш сайт преследуется в соответствии с Российским законодательством об охране авторских прав.
|
| |
|
|
| |
|
***
ЛИТЕРАТУРА
Ссылки из упомянутой статьи (выделено, ADLERZ) с моими дополнениями (например, Kotzias) по роду Formica (род Formica упоминается в 4597 статьях, в том
числе в заголовках 1868 статей, вид Formica picea - в 52, вид Formica candida - в 2 ссылках, использованной здесь базы Formis-2003):
-
АРНОЛЬДИ К.В. ДЛУССКИЙ Г.М. 1978: Надсем. Formicoidea.
1. Сем. Formicidae - муравьи. In: Медведев Г.М. (ред.): Определитель насекомых Европ.ч.СССР. Том 3. Перепончатокрылые, первая часть. – Определители к
фауне СССР. 119: 519-556.
Определитель муравьев, включая упомянутых в этом
обзоре.
-
Берман Д. И., Жигульская З. А., Лейрих А. Н. 1981. Распространение, особенности экологии и зимовки Formica picea Nyl. и Myrmica sp. 2
(Formicidae) на Северо-Востоке СССР // Проблемы почв. зоологии: Тез. докл. VII Всес. сов. Киев, 1981. С. 27 – 28.
-
Берман Д. И., Жигульская З. А., Лейрих А. Н. 1987. Зимняя экология полярного муравья (Formica gagatoides) в верховьях Колымы // Зоол. журн.
1987. Т. 66, № 3. С. 373 – 384.
-
ДЛУССКИЙ Г.М. 1967: Муравьи рода Formica. "Наука", Москва, 236 стр.
Самый
полный обзор систематики, фауны и биологии муравьев этого рода, включая упомянутых в нашем обзоре.
-
Жигульская З. А. 1966. Муравьи Formica picea как фактор почвообразования в каштановых и солонцово-солончаковых почвах Юго-Восточного Забайкалья
// Проблемы почв. зоологии: Тез. докл. II Всес. сов. М., 1966. С. 54 – 55.
-
Жигульская З. А. 1975. Особенности гнездостроения Formica picea Nyl. в вертикально-поясных ландшафтах аридных котловин Тувы // Роль животных в
формировании экосистем. М., 1975. С. 82 – 84.
-
НАСОНОВ Н.В. 1889: Материалы по естественной истории муравьев преимущественно России. – Труды Лаборатории Зоологического Музея Императорского
Музея Московского Университета. 58: 1-78.
-
ADLERZ, G. 1914: Formica fusca picea NYL., en torfmossarnas
myra. – Arkiv fur Zoologi 8 (No 26): 1-5.
-
BOLTON, B. 1995: A New General Catalogue of the Ants
of the World. – Harvard University Press, Cambridge,
Massachusetts and London, 504 pp.
-
BONNER, W. 1915: Die Uberwinterung von Formica picea
und andere biologische Beobachtungen. – Biologisches
Zentralblatt 35: 65-77.
-
Czechowska, W. and W. Czechowski (1998). "Species of ants (Hymenoptera, Formicidae) new to the Sandomierska Lowland. (in Polish)." Fragm. Faun.
(Warsaw) 41: 251-254.
-
COLLINGWOOD, C.A. 1979: The Formicidae of Fennoscandia
and Denmark. – Fauna Entomologica Scandinavica
8: 1-174.
-
DE LATTIN, G. 1967: Grundriss der Zoogeographie. –
Fischer, Jena, 602 pp.
-
DLUSSKY,G. & PISARSKI, B. 1971: Rewiizja polskich gatunkow
mrowek (Hymenoptera:Formicidae) z rodzaju
Formica L. – Fragmenta Faunistica 16: 145-224.
-
EMERY, C. 1925: Hymenoptera. Fam. Formicidae. Subfam.
Formicinae. – Genera Insectorum 183: 1-302
-
GOROPASHNAYA, A. 2003: Phylogeographic structure and
genetic differentiation in Formica Ants. Acta Universitatis
Upsaliensis. – Comprehensive Summaries of
Uppsala Dissertations from the Faculty of Science and
Technology 912: 1-36.
-
GOROPASHNAYA, A., FEDOROV, V.B., SEIFERT, B. &
PAMILO, P. 2004a: Limited phylogeographic structure
across Eurasia in two red wood ant species Formica
pratensis and F. lugubris (Hymenoptera, Formicidae).
– Molecular Ecology 13: 1849-1858.
-
GOROPASHNAYA, A., FEDOROV, V.B. & PAMILO, P. 2004b:
Recent speciation in the Formica rufa group ants (Hymenoptera:
Formicidae): inference from mitochondrial
DNA phylogeny. – Molecular Phylogenetics and Evolution
32: 198-206.
-
GYLLENBERG, G. & ROSENGREN, R. 1984: The oxygen consumption
of submerged Formica queens (Hymenoptera,
Formicidae) as related to habitat and hydrochoric
transport. – Annales Entomologici Fennici 50: 76-80.
-
Kotzias, H. (1930). "Formica picea Nyl. in Schlesien." Zool. Anz. 92: 56-58.
-
KUTTER, H. 1917: Myrmecologische Beobachtungen. –
Biologisches Zentralblatt 37: 429-437.
-
KUTTER, H. 1977: Hymenoptera-Formicidae. In: SAUTER,
W. (Ed.): Fauna Insecta Helvetica 6. – Schweizerische
Entomologische Gesellschaft Zurich, Zurich, 298 pp.
-
NYLANDER, W. 1846: Adnotationes in monographiam
formicarum borealium Europae. – Acta Societaris Scientiarum
Fennicae 2: 875-944.
-
Roszler, P. (1937). "Beitrag zur Kenntnis der Verbreitung der Serviformica picea Nyl." Entomol. Rundsch. 55: 57-60.
-
SEIFERT, B. 2002: How to distinguish most similar insect
species - improving the stereomicroscopic and mathematical
evaluation of external characters by example of
ants. – The Journal of Applied Entomology 126: 445-454.
-
SMITH, F. 1878: Scientific Results of the Second Yarkand
Mission; based upon the collections and notes of the late
Ferdinand Stoliczka, Ph.D. Hymenoptera. – Superintendent
of Government Printing (Government of India),
Calcutta, 22 pp.
-
SORENSEN, U. 2001: Untersuchung zur Variabilitat der
Neststandorte von Coptoformica forsslundi LOHMANDER,
1949 (Hymenoptera:Formicidae) im Naturschutzgebiet
"Suderlugumer Binnendunen" (Nordfriesland).
– Myrmecologische Nachrichten 4: 17-24.
-
YARROW, I.H.H. 1954: The British ants allied to Formica
fusca L. – Transactions of the Society for British Entomology
11: 229-224.
|
| |
|
©2005, Vladislav
Krasilnikov (translation & supplement)
|
|
Всякое использование без согласования с автором и без активной гиперссылки на наш сайт
преследуется в соответствии с Российским законодательством об охране авторских прав.
|
|
